Головной и спинной мозг
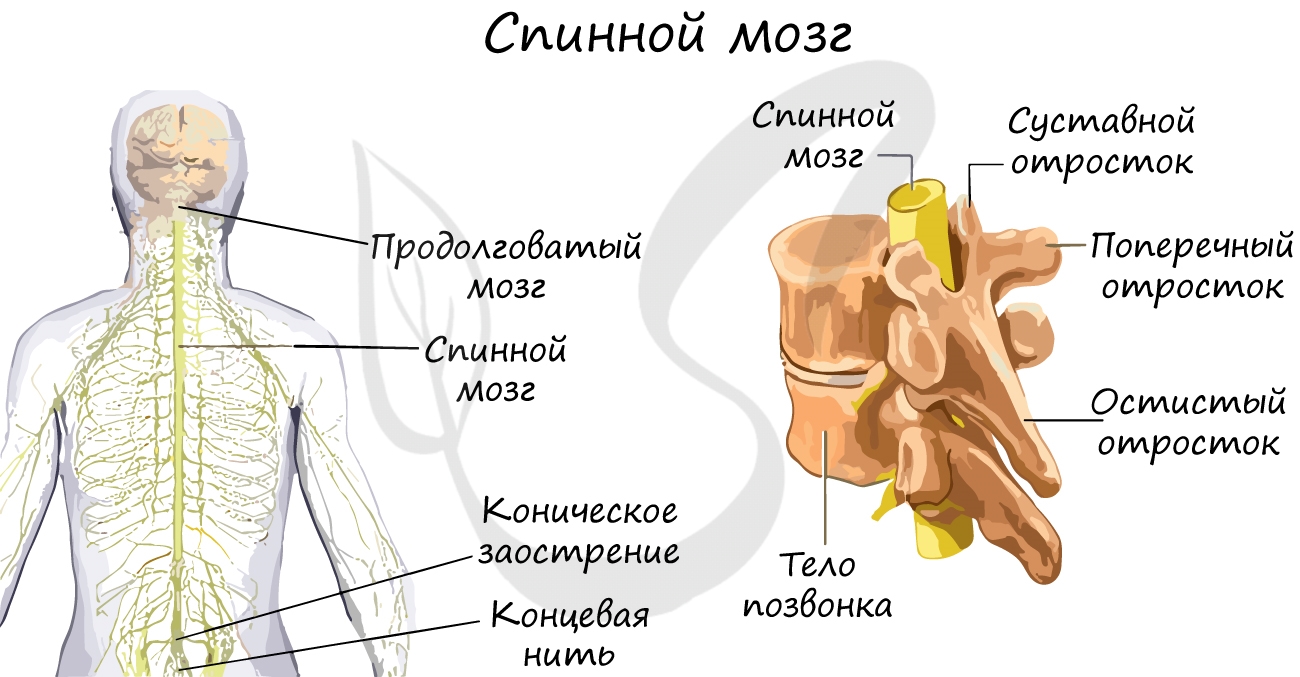
Спинной мозг
Представляет собой нервный тяж, лежащий в образованном позвонками позвоночном канале. Тянется от затылочного отверстия до поясничного отдела позвоночника. Вверху переходит в продолговатый мозг, внизу заканчивается коническим заострением с концевой нитью.
Спинной мозг покрыт несколькими оболочками: твердой мозговой, паутинной и мягкой. Между паутинной и мягкой оболочками циркулирует спинномозговая жидкость — ликвор, окружающая спинной мозг и принимающая активное участие в обмене веществ спинного мозга.
На поперечном срезе спинной мозг (СМ) напоминает бабочку. В центре расположено серое вещество, состоящее из тел нейронов. На периферии расположено белое вещество, которое образовано отростками нейронов.
В сером веществе СМ различают два передних выступа (передние рога), два боковых (боковые рога) и два задних (задние рога). В следующей статье мы будем изучать рефлекторные дуги, так что эти знания нам очень пригодятся. В рогах серого вещества находятся нейроны, которые входят в состав рефлекторных дуг.
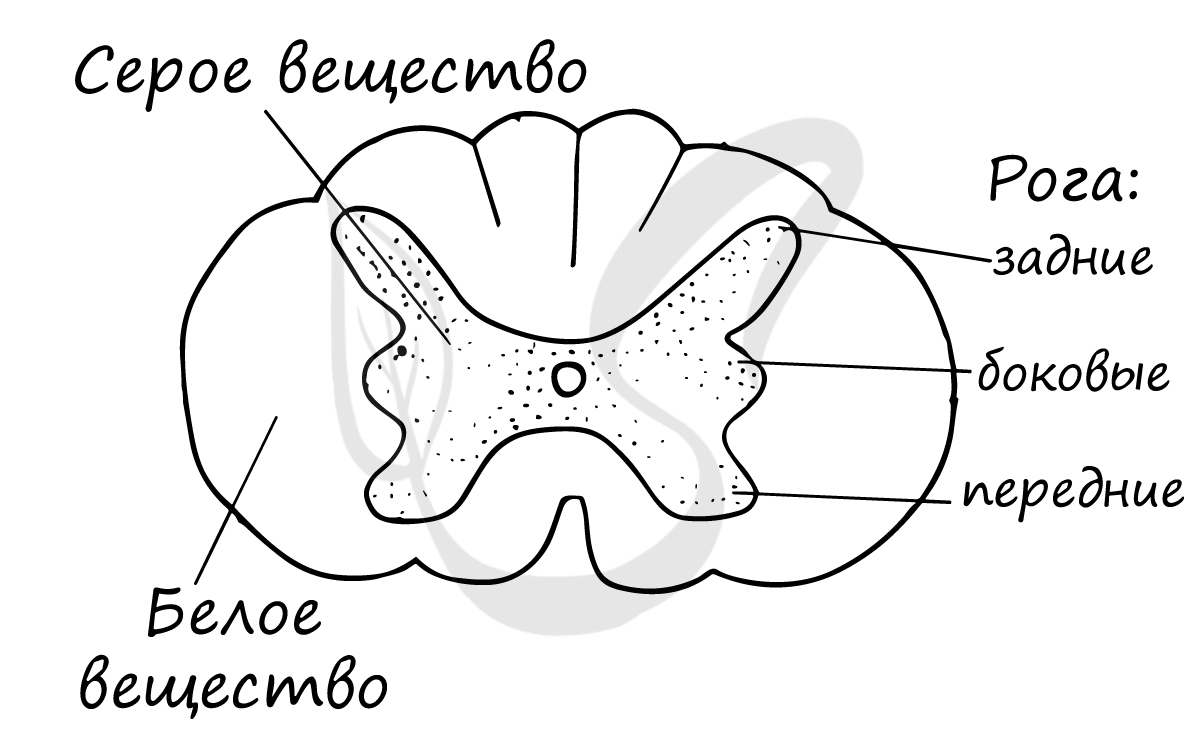
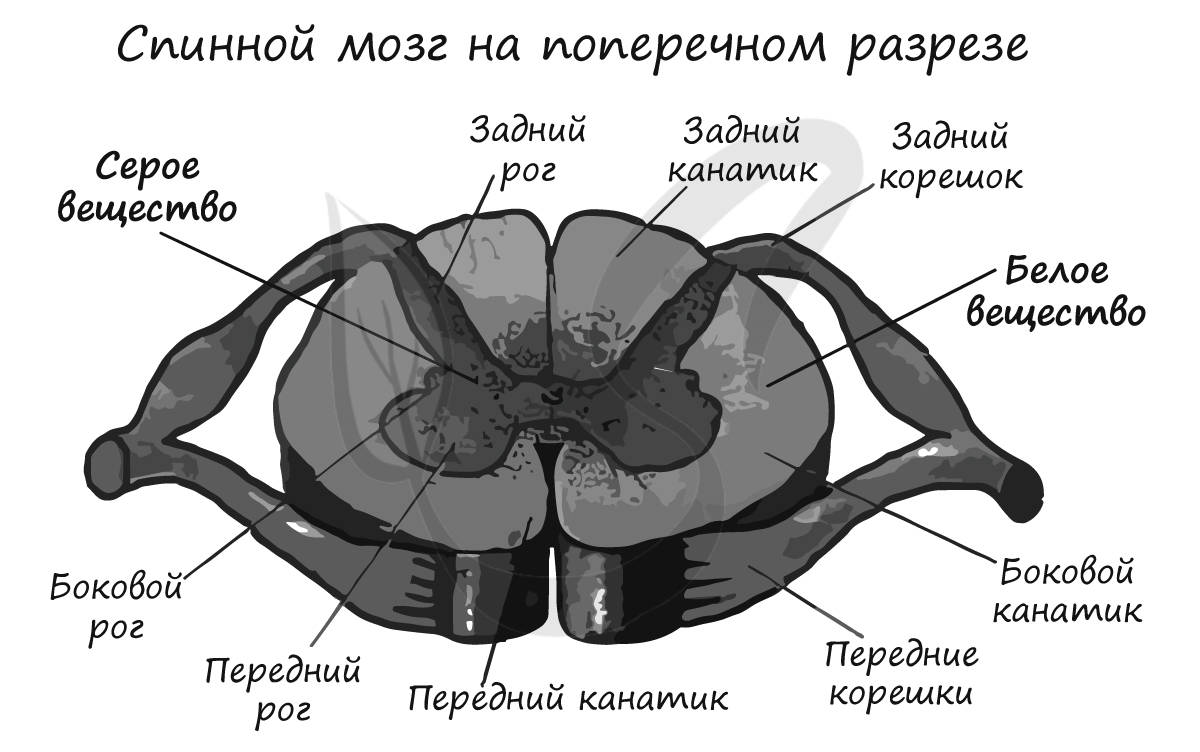
К задним рогам спинного мозга подходят многочисленные нервные волокна, которые, объединяясь, образуют пучки — задние корешки. Из передних рогов спинного мозга выходят многочисленные нервные волокна, которые образуют — передние корешки.
Белое вещество состоит из многочисленных нервных волокон, пучки которых образуют канатики. Пути спинного мозга подразделяются на восходящие — от рецепторов к головному мозгу, и нисходящие — от головного мозга к органам-эффекторам. От спинного мозга отходит 31 пара спинномозговых нервов.
У спинного мозга выделяют две важнейшие функции:
- Рефлекторную
За счет тел нейронов, которые расположены в сером веществе спинного мозга и входят в состав рефлекторных дуг, обеспечивающих рефлексы.
За счет наличия в спинном мозге белого вещества, в состав которого входят многочисленные нервные волокна, образующие пучки и канатики вокруг серого вещества.
Головной мозг и его отделы
Мы переходим к изучению головного мозга человека, сложноустроенного главного органа центральной нервной системы, расположенного в надежном костном вместилище — черепе. Масса мозга в среднем составляет от 1300 до 1500 грамм.
Замечу, что вес мозга никак не связан с интеллектуальными способностями: так у Альберта Эйнштейна головной мозг весил 1230 грамм — меньше, чем у среднестатистического человека. Интеллект скорее определяется сложностью и разветвленностью нейронных сетей мозга, но никак не массой.

В мозге человека выделяют пять отделов: продолговатый, задний (мост и мозжечок), средний, промежуточный и конечный. Наиболее древние отделы — продолговатый, задний и средний — образуют ствол мозга, напоминающий по строению спинной мозг. Иногда к стволу мозга относят и промежуточный отдел. От ствола мозга отходят 12 пар черепных нервов.
Конечный мозг отличается от строения ствола мозга, он представляет собой огромное скопление (около 16 млрд.) нейронов, которые образуют кору больших полушарий (КБП). Нейроны располагаются в несколько слоев, их отростки образуют тысячи синапсов с другими нейронами и их отростками. В КБП расположены центры высшей нервной деятельности — памяти, мышления, речи.
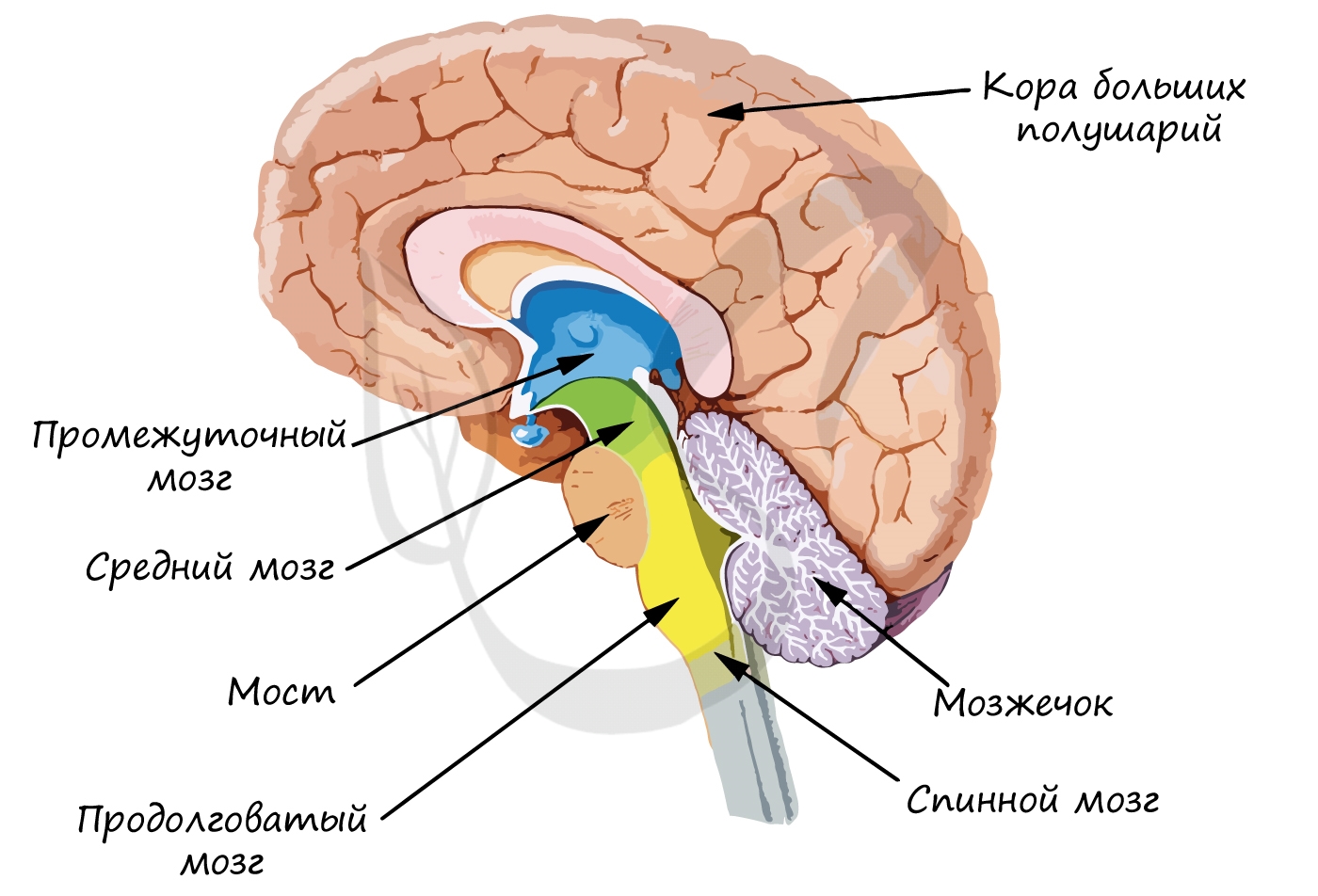
Мы начинаем увлекательное путешествие по отделам головного мозга. Для вас принципиально важно разделить между собой и запомнить функции различных отделов, для этого обязательно используйте воображение!)
Самый древний отдел головного мозга. Запомните, что он регулирует жизненно важные функции: сердечно-сосудистую систему, процессы дыхания и пищеварения. Здесь сосредоточены центры защитных рефлексов — рвоты, чихания, кашля.
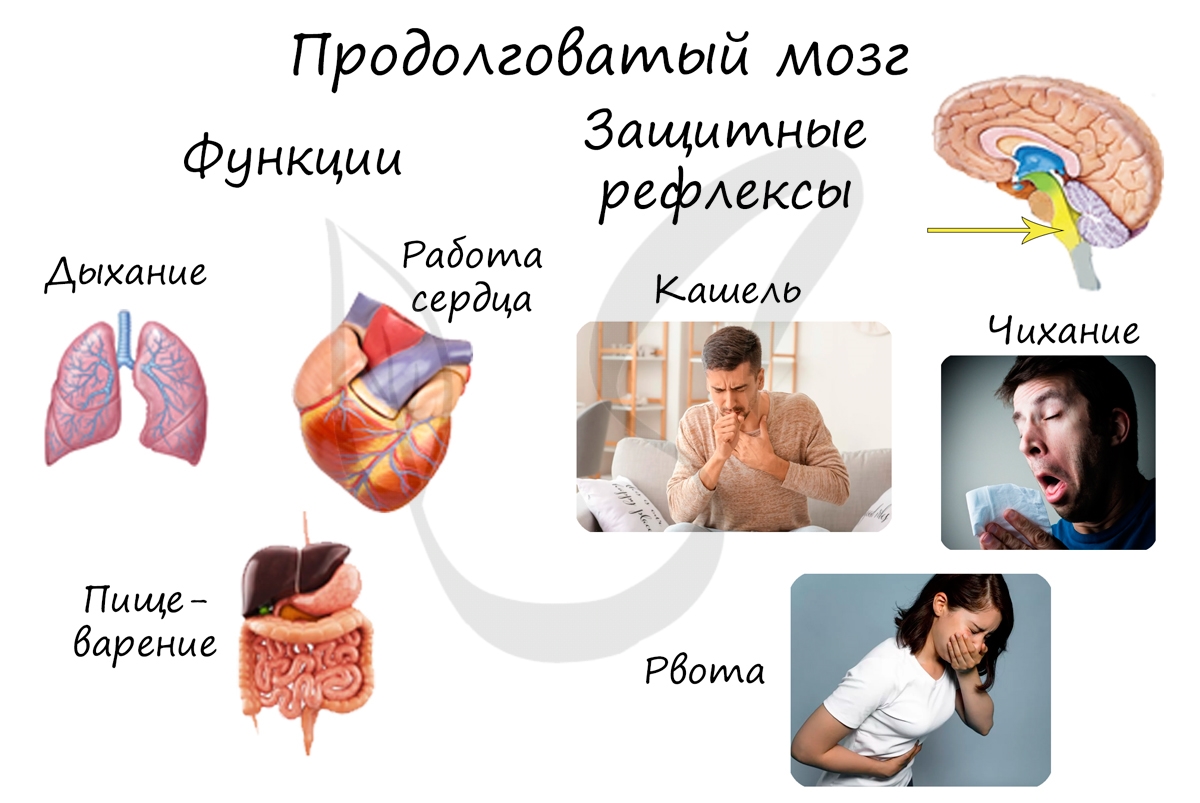
Варолиев мост выполняет проводниковую функцию: через мост проходят все нисходящие и восходящие нервные пути. Также он контролирует работу мимических и жевательных мышц лица, слезной железы.
Мозжечок имеет свои собственные полушария, соединенные друг с другом. Кора мозжечка образована серым веществом, подкорковые ядра окружены белым веществом.
Мозжечок принимает участие в координации произвольных движений, способствует сохранению положения тела в пространстве, регулирует тонус и равновесие. Благодаря мозжечку наши движения четкие и плавные.

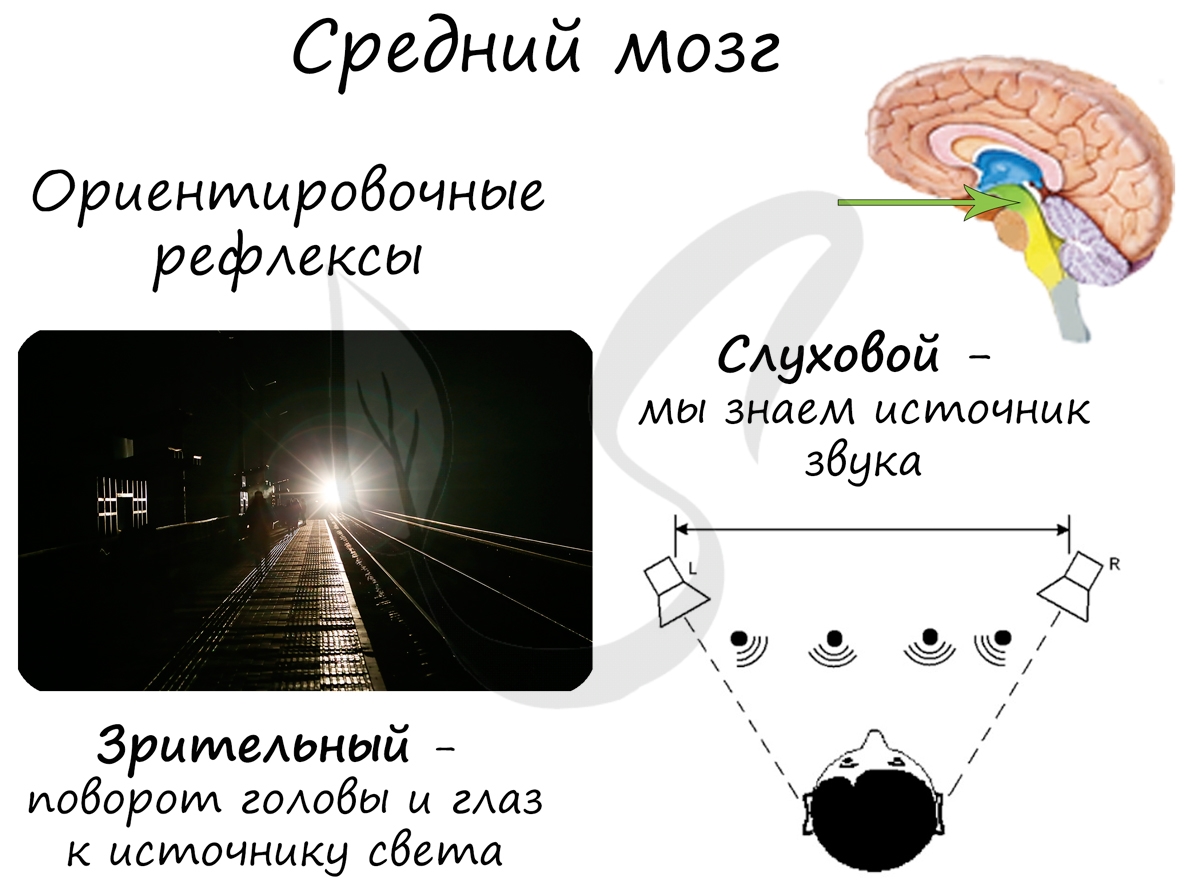
В среднем мозге находятся верхние (передние) и нижние (задние) бугры четверохолмия. Верхние бугры четверохолмия отвечают за зрительный ориентировочный рефлекс, а нижние — за слуховой ориентировочный рефлекс.
В чем выражается зрительный ориентировочный рефлекс? Представьте, что заходите в темную комнату. В ее уголке уютно сияет экран, виден сайт (конечно же) студариум =) И тут начинается зрительный ориентировочный рефлекс: Вы двигаете глазами, поворачиваете голову в направлении источника интеллектуального света. Не забываете при этом регулировать величину зрачка и аккомодацию глаз — все это зрительный ориентировочный рефлекс.
Слуховой ориентировочный рефлекс также необходим для нас. Хорошо, если, читая учебник сейчас, вы находитесь в тишине. Вдруг у вас начинает звонить телефон: вы тотчас перестаете читать и направляетесь к источнику звука — телефону. Благодаря этому ориентировочному рефлексу мы можем определять место источника звука относительно нас (слева, справа, сзади, спереди).
Средний мозг также выполняет проводниковую функцию, участвует в регуляции мышечного тонуса и позы тела.
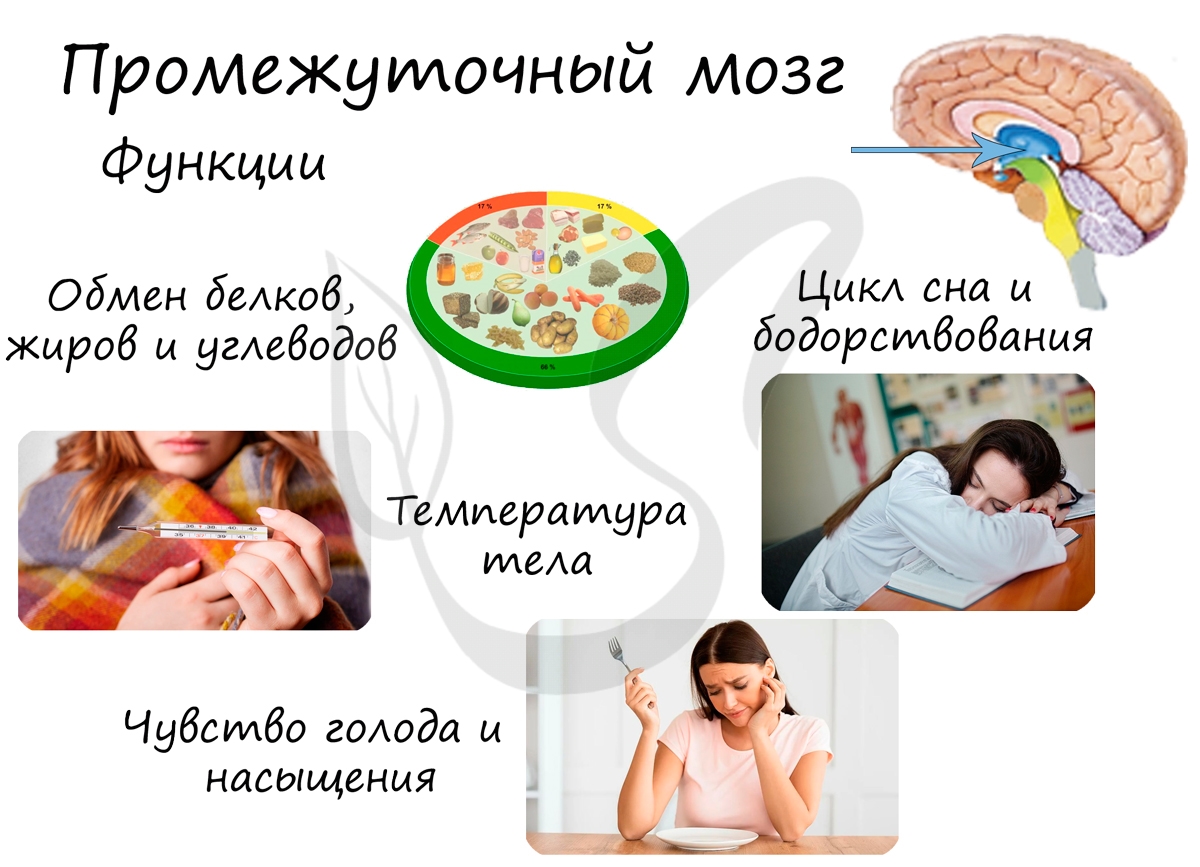
Напомню, что изученный нами гипоталамус, связанный с ним гипофиз, эпифиз и таламус относятся к промежуточному мозгу. Вам известно, что гипоталамус руководит гипофизом — дирижером желез внутренней секреции, поэтому функциями гипоталамуса являются: регуляция обмена белков, жиров и углеводов, а также водно-солевой обмен.
Помимо этого, гипоталамус контролирует симпатическую и парасимпатическую системы, регулирует температуру тела, отвечает за циклы сна и бодрствования. В гипоталамусе находятся центры голода и насыщения.
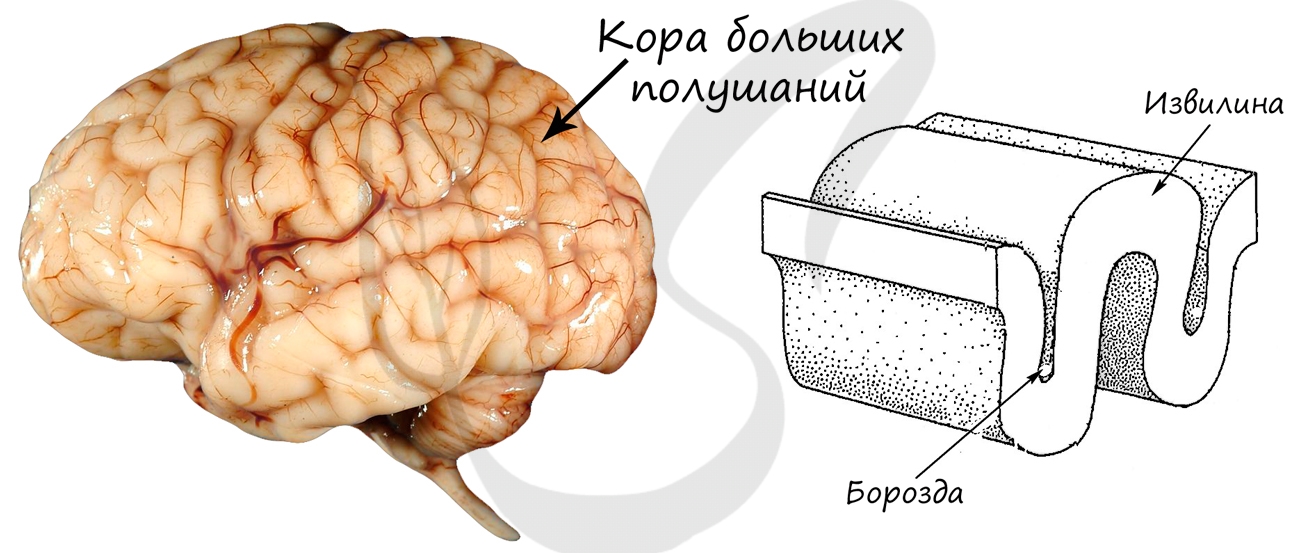
Состоит из подкорковых структур и коры больших полушарий (КБП). Поверхность КБП достигает в среднем 1,5-1,7 м 2 . Такая большая площадь обусловлена тем, что КБП образует извилины — возвышения мозгового вещества, и борозды — углубления между извилинами.
Кора больших полушарий
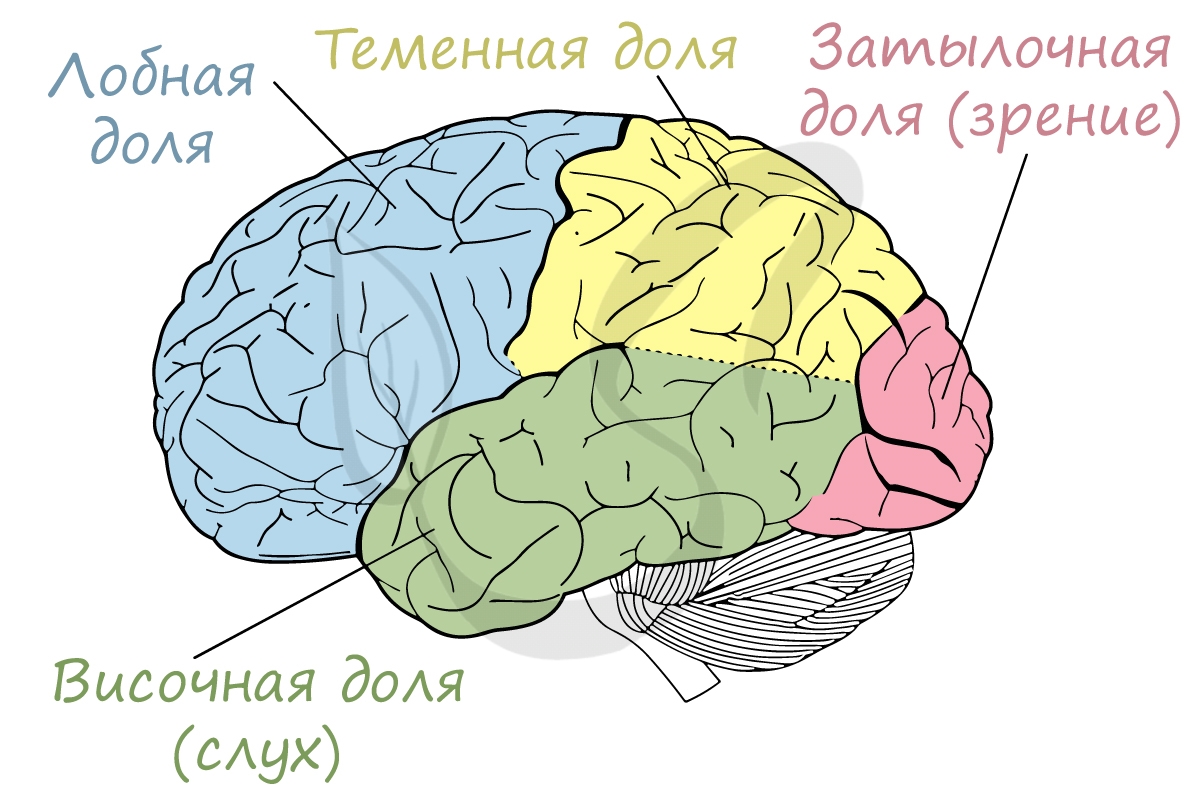
В коре имеется несколько слоев клеток, между которыми образуются многочисленные разветвленные связи. Несмотря на то, что кора функционирует как единый механизм, разные ее участки анализируют информацию от разных периферических рецепторов, которые И.П. Павлов называл корковыми концами анализаторов.
Корковое представительство зрительного анализатора располагается в затылочной доле КБП, именно в связи с этим при падении на затылок человек видит «искры из глаз», когда нейроны этой доли возбуждаются механически, вследствие удара.
Корковое представительство слухового анализатора находится в височной доле коры больших полушарий.
Запомните, что корковое представительство двигательного анализатора — моторная зона — находится в передней центральной (прецентральной) извилине, а представительство кожного анализатора — сенсорная зона — в задней центральной (постцентральной) извилине.
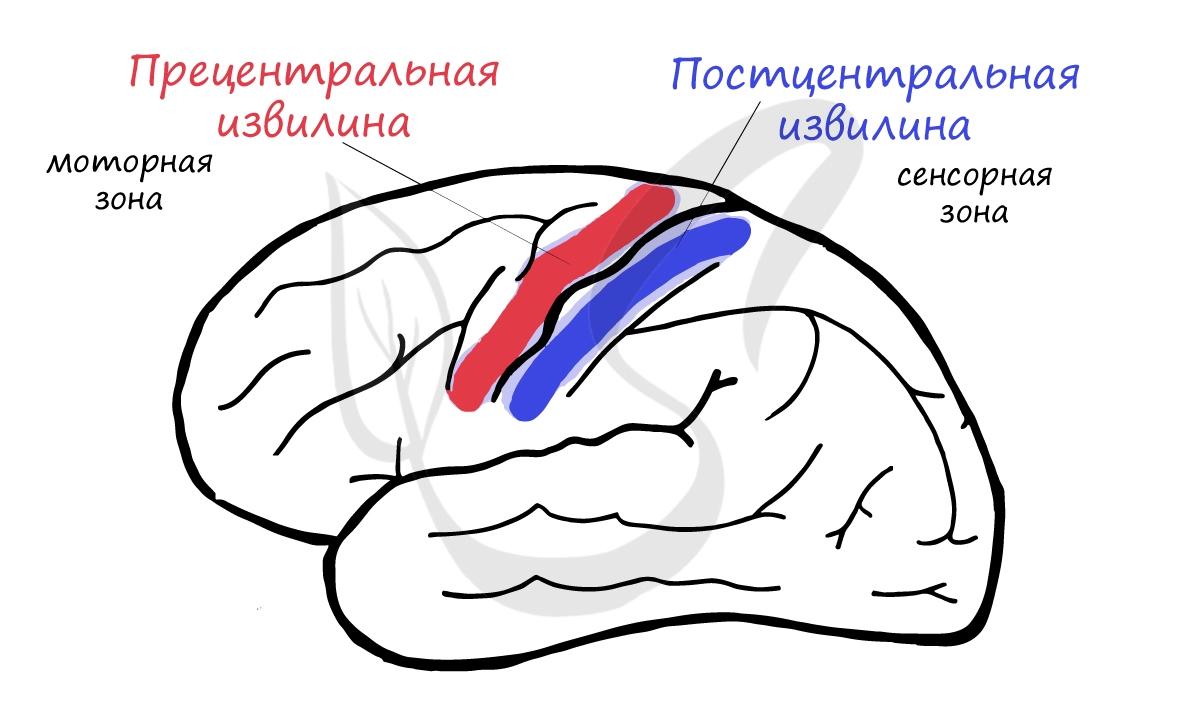
Вдумайтесь! При совершении любого произвольного (осознанного) движения нервный импульс возникает именно в нейронах прецентральной извилины, откуда начинает свой длинный путь через ствол мозга, спинной мозг и, наконец, достигает органа-эффектора.
Импульсы от кожных рецепторов достигают нейронов постцентральной извилины — сенсорного отдела, благодаря чему мы получаем от них информацию и осознаем собственные ощущения.
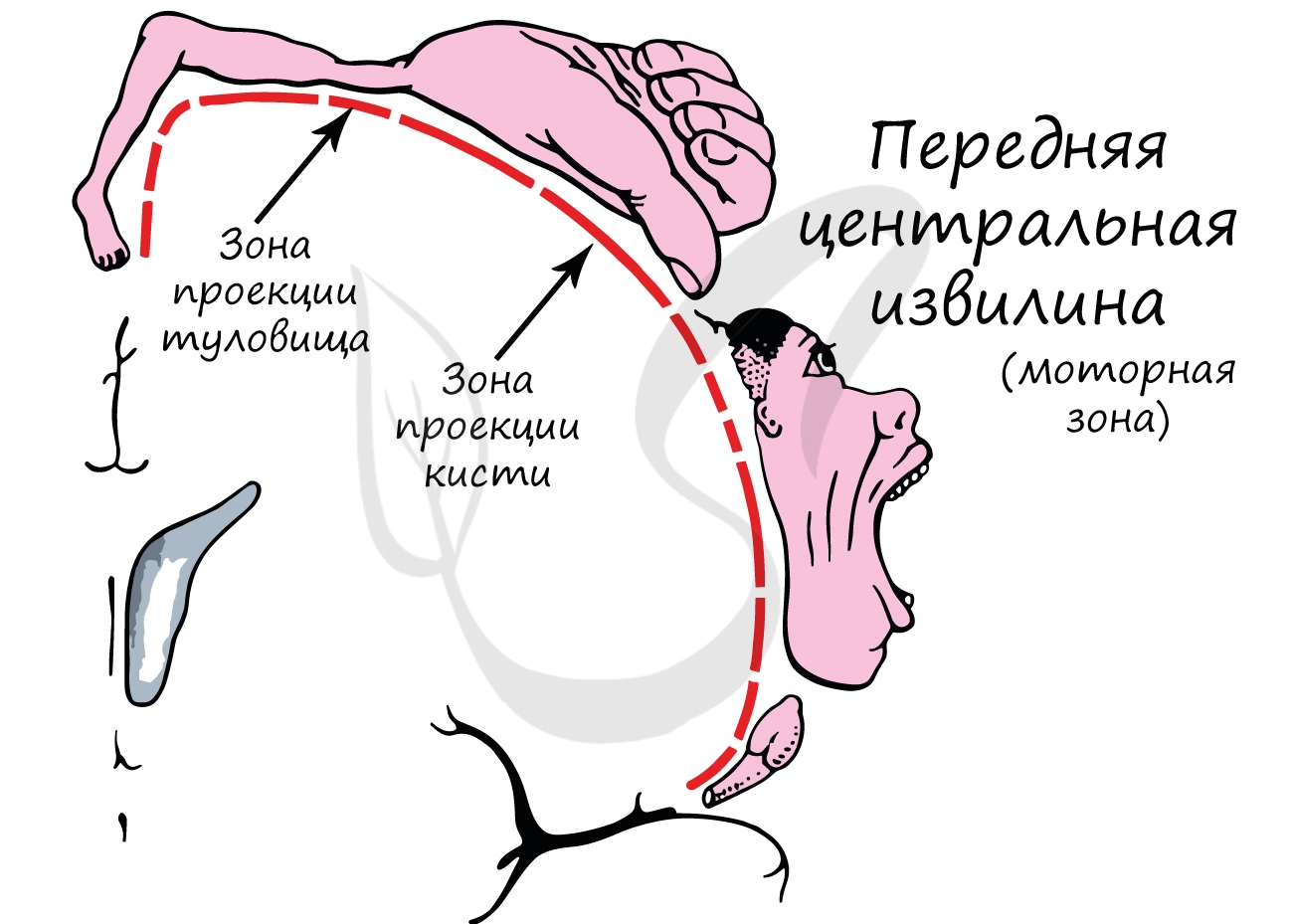
Количество нейронов в этих извилинах, отведенных для различных органов, неодинаково. Так зона проекции пальцев кисти занимает много места, благодаря чему становятся возможны тонкие движения пальцами. Зона проекции мышц туловища гораздо меньше зоны пальцев, так как движения туловища более однообразные и менее сложные.
Изученные нами участки мозга, в которых происходит преобразование и анализ поступающей информации, называются ассоциативными зонами КБП. Эти зоны связывают различные участки КБП, координируют ее работу, играют важнейшую роль в образовании условных рефлексов.
Наша осознанная деятельность лежит в рамках коры больших полушарий: любое осознанное движение, любое ощущение (температурное, болевое, тактильное) — все имеет представительства в КБП. Кора — основа связи с внешней средой, адаптации к ней. В фундаменте процесса мышления также лежит КБП. В общем, вы поняли, как высоко надо ее ценить и как хорошо знать данную тему 🙂
Вы наверняка слышали, что функционально правое и левое полушария отличаются. В левом полушарии находятся механизмы абстрактного мышления (языковые способности, аналитическое мышление, логика), а в правом — конкретно-образного (воображение, параллельная обработка информации). При травмах, повреждениях левого полушария может нарушаться речь.

Заболевания
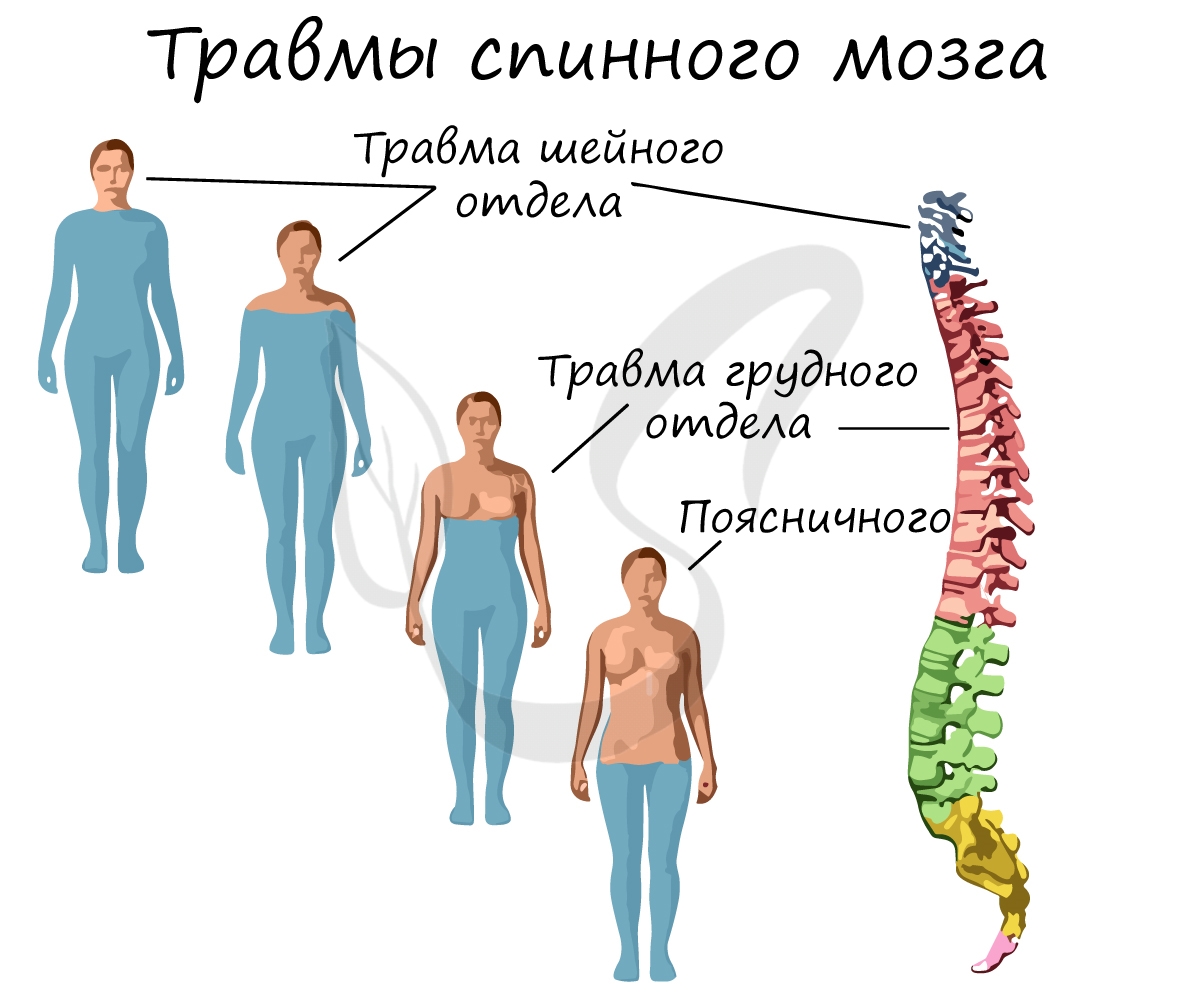
В зависимости от уровня поражения спинного мозга при травме картина неврологических нарушений проявляется по-разному. Чем выше уровень поражения, тем больше нервных путей оказываются «отрезанными» от головного мозга. Так, к примеру, при травме поясничного отдела движения руками сохранены, а при травме шейного — движения руками невозможны.
Иногда после инсульта (кровоизлияния в ткани мозга) или травмы развивается паралич (полное отсутствие движений) на одной из сторон тела. Зная анатомию, вы можете седлать вывод: если движения пропали в правой руке и ноге, то инсульт произошел слева.
Почему существует такая закономерность? Дело в том, что нервные волокна, идущие от прецентральной извилины к рабочим органам — мышцам, формируют так называемый физиологический перекрест на границе продолговатого и спинного мозга. То есть, говоря проще: часть нервов, которые шли от левого полушария переходят на правую сторону и наоборот — нервы от правого полушария переходят на левую сторону.
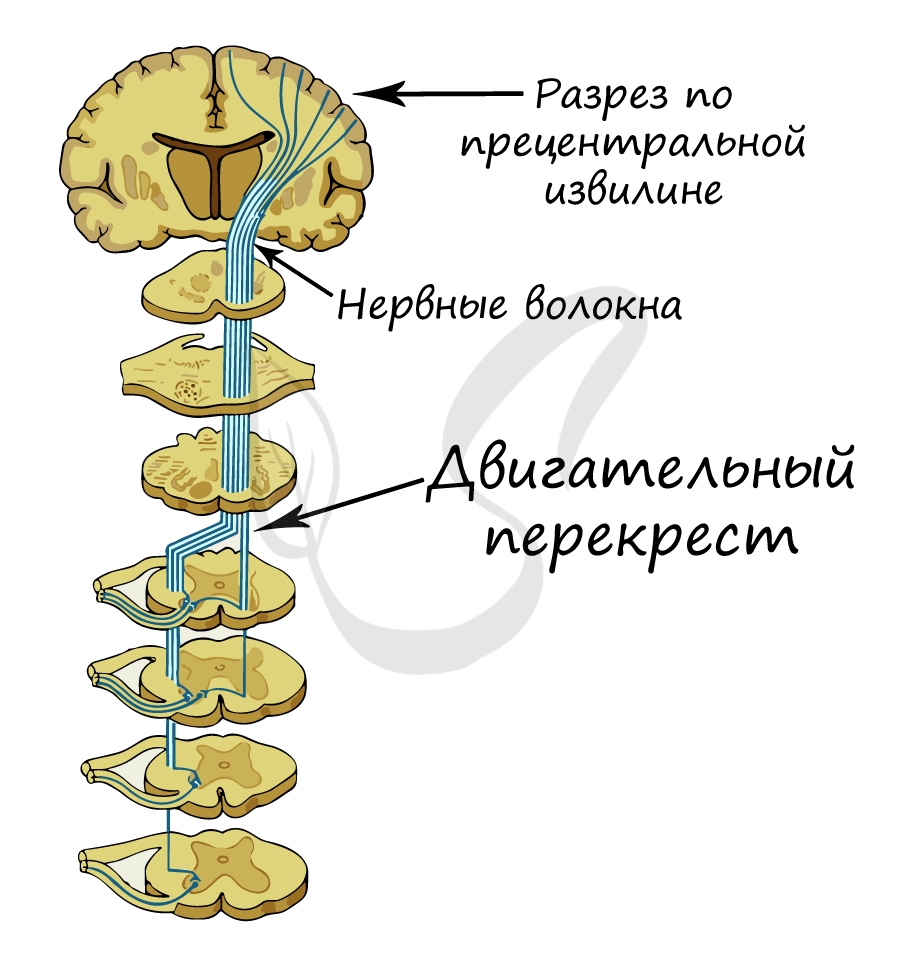
© Беллевич Юрий Сергеевич 2018-2021
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Источник
Зоны мозга контролирующие мышечные чувства
а) Первичная моторная кора. Первичная моторная кора (поле 4) — полоска агранулярной коры в прецентральной извилине. Она дает начало 60-80% (количество варьирует) волокон корково-спинномозгового пути (КСП). Остальная часть волокон отходит от премоторной, поясной и добавочной моторной зон, а также теменной коры, что показано в отдельной статье на сайте. Наибольшую плотность окончаний КСП в спинном мозге наблюдают в зонах, иннервирующих дистальные мышцы конечностей.
Части тела противоположной стороны соматотопически представлены в коре инвертированными, за исключением лица, а относительно крупные области занимают кисть (играет важную роль в контроле движений пальцев), околоротовая область и язык. Зону кисти обычно можно определить по направленному кзади возвышению в 6-7 см от верхнего края полушария.
Ипсилатералъные части тела также имеют соматотопическую организацию; ипсилатеральные моторные нейроны составляют лишь 10% волокон пирамидного пути, которые не переходят на противоположную сторону и не иннервируют дистальные мышцы конечностей.
При целенаправленной стимуляции моторной коры человека установлено, что клеточные колонны регулируют направление движения. Первичная моторная кора «синтезирует» двигательные команды, но не является местом их возникновения. От нее волокна в составе КСП направляются в спинной мозг и широко разветвляются по мере достижения места своего окончания. Для того чтобы взять, например, ручку в руку, требуется:
(а) умеренное сокращение мышцы, противопоставляющей большой палец, как основной движущей мышцы;
(б) сокращение с той же интенсивностью отдела глубокого сгибателя пальцев, от которого отходит сухожилие к дистальной фаланге указательного пальца;
(в) более слабое сокращение мышцы, приводящей большой палец, и
(г) короткого сгибателя большого пальца.
Удержание всей руки при любом типе манипуляций — функция премоторной коры, отражающая важность бессознательной регуляции положения тела при совершении произвольных движений. Более крупные моторные зоны коры формируются при «объединении» близлежащих нейронных колонн на основе их функций и создании сложных последовательностей движений.
1. Пластичность моторной коры. Некрупные поражения моторной коры у обезьян и низших млекопитающих приводят первоначально к параличу соответствующей части тела, а затем в течение нескольких дней (иногда часов) — к быстро прогрессирующему восстановлению функции. Восстановление может быть обусловлено изменением свойств клеточных колонн, расположенных рядом с местом поражения, которые берут на себя утраченную двигательную функцию. Увеличение двигательной зоны участка коры возможно путем локального введения антагонистов ГАМК в кору. Расширение моторной зоны на уровне спинного мозга заранее обеспечено обширными зонами перекрытия от поля 4 до колонок двигательных нейронов серого вещества переднего рога, однако степень пластичности здесь ниже, чем в коре.
Взаимосвязи между КСП (и другими нисходящими путями) и двигательными нейронами в спинном мозге возникают посредством вставочных нейронов. Во вставочных нейронах происходит объединение сенсорной и корковой информации, что сопровождается специфической и организованной активацией пулов двигательных нейронов и сокращением иннервируемых ими мышц.
2. Источники афферентных путей к первичной моторной коре:
— Моторная кора противоположной стороны через мозолистое тело. Наиболее прочные комиссуральные связи возникают между соответствующими колоннами клеток, иннервирующими мышцы живота и спины. Это вполне понятно, поскольку данные группы мышц обычно сокращаются одновременно с двух сторон, поддерживая вертикальное положение головы и туловища. Самые слабые комиссуральные связи имеются между колоннами клеток, контролирующими дистальные группы мышц конечностей, движения в которых слева и справа происходят независимо друг от друга.
— Соматосенсорная кора. Кожные колонны клеток в полях 1, 2 и 3 отдают кпереди короткие ассоциативные волокна (взаимосвязи с зоной кисти особенно многочисленные; эти волокна короткие, так как зоны, отвечающие за кисть в сенсорной и моторной коре, обычно занимают соседние стенки центральной борозды). Проприоцептивные клеточные колонны получают афферентые волокна от аннуло-спиральных окончаний мышечных веретен; они отдают короткие ассоциативные волокна к соответствующим двигательным колоннам, активируя рефлексы растяжения с длинной дугой.
— Противоположное зубчатое ядро. Мозжечок принимает участие в выборе нужных мышц-синергистов, а также времени и силы их сокращений.
— Дополнительная моторная зона (ДМЗ).
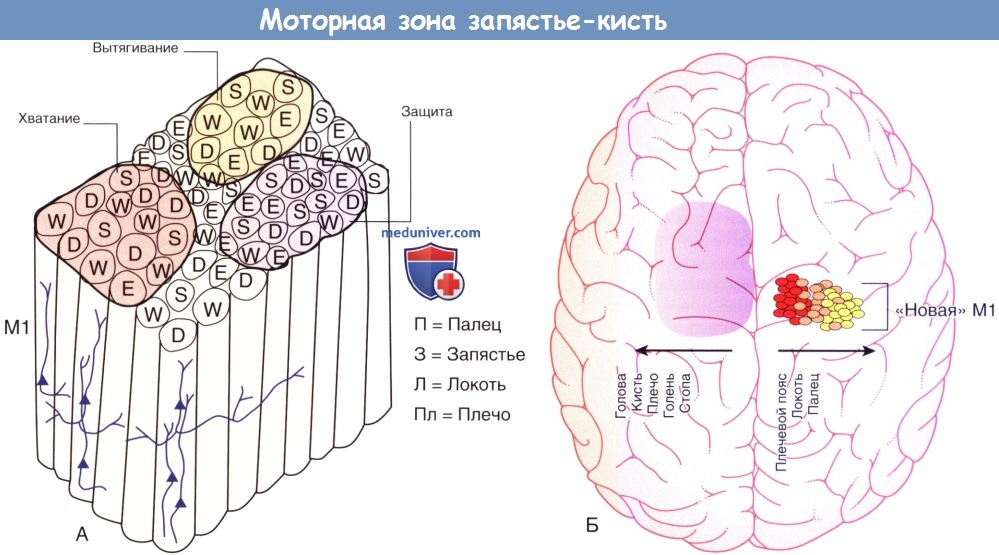 (А) Предполагаемая функциональная организация сегмента запястье-кисть в первичной двигательной коре (М1) у обезьян и других приматов.
(А) Предполагаемая функциональная организация сегмента запястье-кисть в первичной двигательной коре (М1) у обезьян и других приматов.
Несмотря на то, что М1 имеет в целом соматотопическую организацию, локальная соматотопическая структура разбита в виде мозаики на радиальные ряды нейронов, инициирующих небольшие специфические движения.
Миниколонны, отвечающие за движения отдельных пальцев, могут примыкать к колоннам запястья, локтя или плеча, а подгруппы этих миниколонн объединены по функции образования более сложных последовательностей движений, таких как хватание, вытягивание руки или защита рукой от удара.
(Б) Пространственная организация моторных нейронов коры приматов, контролирующих движения мышечных групп.
От медиальной стороны к латеральной они располагаются в следующей последовательности: стопа, голень, плечо, кисть и голова (голубой).
В каудальном отделе М1 лежат двигательные нейроны, взаимодействующие непосредственно с двигательными нейронами (спинного мозга) и отвечающие за высокоточные движения, необходимые для выполнения тонких двигательных навыков.
Эти нейроны расположены в медиально-латеральной последовательности от проксимальных (красный) до дистальных (желтый) групп мышц. Авторы обозначают эту зону как «новую М1», подотдел «старой М1» (голубая) для того, чтобы отразить недавнее возникновение сложной двигательной активности и эволюционно «нового» отдела моторной коры.
б) Премоторная кора. Премоторная кора (поле 6 на латеральной поверхности полушария) примерно в 6 раз крупнее первичной моторной коры. Она получает когнитивные импульсы от лобной доли при намерении произвести движение, а также от теменной доли (поле 7), передающей огромное количество тактильной и пространственно-зрительной информации. Максимальная ее активация происходит, когда последовательность движений происходит в ответ на зрительные или соматосенсорные раздражители, такие как приближение к объекту, полностью находящему в поле зрения, или идентификация объекта, находящегося вне поля зрения путем ощупывания. Активация премоторной коры, как правило, двусторонняя. Одним из объяснений служит межполушарная передача двигательных программ через мозолистое тело.
Следует также принимать во внимание обширные связи премоторной коры с ядрами ствола мозга, дающими начало ретикуло-спинномозговым путям (и небольшому пути к КСП). Поражения, затрагивающие премоторную кору у человека, встречают редко, они характеризуются нестабильностью положения в противоположном плече и бедре. Таким образом, важной функцией премоторной коры, вероятно, служит двустороннее поддержание позы, например фиксация плечей при работе двумя руками и стабилизация бедер при ходьбе. Премоторная кора может вносить вклад в восстановление функции при истинной моторной гемиплегии, возникающей вследствие сосудистого поражения, затрагивающего КСП в лучистом венце. При данном поражении во время ПЭТ-исследования наблюдают повышение активности премоторной коры; кпереди от КСП спускается корково-ретикуло-спинномозговой проводящий путь.
в) Дополнительная моторная зона (ДМЗ). В отличие от премоторной коры, реагирующей на внешние раздражители, ДМЗ (поле 6 на медиальной поверхности полушария) отвечает на внутренние раздражители и принимает особое участие в планировании движений. Это подтверждает тот факт, что ДМЗ возбуждается лобной долей (ДЛПФК) в тот момент, когда мы собираемся совершить движение, даже если движение в итоге не возникает. Важнейшей функцией ДМЗ, вероятно, является предварительное программирование последовательностей движений, уже заложенных в двигательной памяти. ДМЗ функционирует совместно с моторной петлей, проходящей через базальные ганглии, а также взаимодействует с полем 4 и отдает волокна непосредственно в КСП.
Одностороннее поражение ДМЗ может сопровождаться акинезией (невозможностью начать движение) руки и ноги с разных сторон. Двустороннее поражение приводит к тотальной акинезии, включая речевую акинезию.
1. Корковые глазодвигательные поля. На рисунке ниже показаны корковые глазодвигательные поля, участвующие в сканирующих движениях (саккадах). Их взаимодействия и функции представлены в таблице ниже.
2. Дорсолатеральная префронтальная кора (ДЛПФК). ДЛПФК — высший когнитивный центр, участвует в оценке зрительной картины, принятии решения о сознательном совершении саккад и добровольном подавлении рефлекторных саккад (произвольные саккады возникают по воле человека). Рефлекторные саккады — автоматические реакции на объекты, появляющиеся в периферическом поле зрения. Строго говоря, рефлекторные саккады следует называть ответными; они не являются истинными рефлексами, так как способны подавляться волей.
3. Поясная кора. Вместе с ДЛПФК участвует в принятии решений и оценке эмоциональной важности, или ценности, видимых объектов.
4. Дополнительное глазодвигательное поле. Занимает передний отдел ДМЗ и участвует в планировании движений, особенно, если требуются множественные саккадические движения.

5. Фронтальное глазодвигательное поле. Фронтальное глазодвигательное поле (ФГДП) инициирует произвольные саккады, благодаря которым происходит переключение внимания на раздражитель или подавляется стремление к прямому взору на новый раздражитель в ответ на одно или более из трех перечисленных афферентных влияний. ФГДП «поддерживает» карту зрительного пространства относительно глазодвигательных координат и вместе с верхним холмиком играет ключевую роль в создании зрительно-направленных и произвольных саккад; поражения обеих структур приводят к необратимой утрате саккадических движений. Клинические и экспериментальные наблюдения у обезьян показали следующее.
• ФГДП имеют тоническую двустороннюю активность.
• Повышение активности среднего отдела ФГДП с одной стороны приводит к образованию горизонтальных саккад, направленных в сторону противоположного зрительного поля (противоположной саккады).
• Повышение активности верхнего отдела с одной стороны приводит к созданию нисходящей косонаправленной саккады; двусторонняя активация верхнего отдела приводит к обращению взора обоих глаз вертикально вниз.
• Повышение активности в нижнем отделе приводит к аналогичным последствиям относительно взора вверх.
6. Теменное глазодвигательное поле. Теменное глазодвигательное поле (ТГДП) отвечает за создание рефлекторных саккад и побуждает ФГДП к созданию произвольных саккад. ТГДП также участвует в пространственном восприятии, создавая схему зрительной картины.
Поражения префронтальной коры и лобной доли описаны в отдельной статье на сайте.
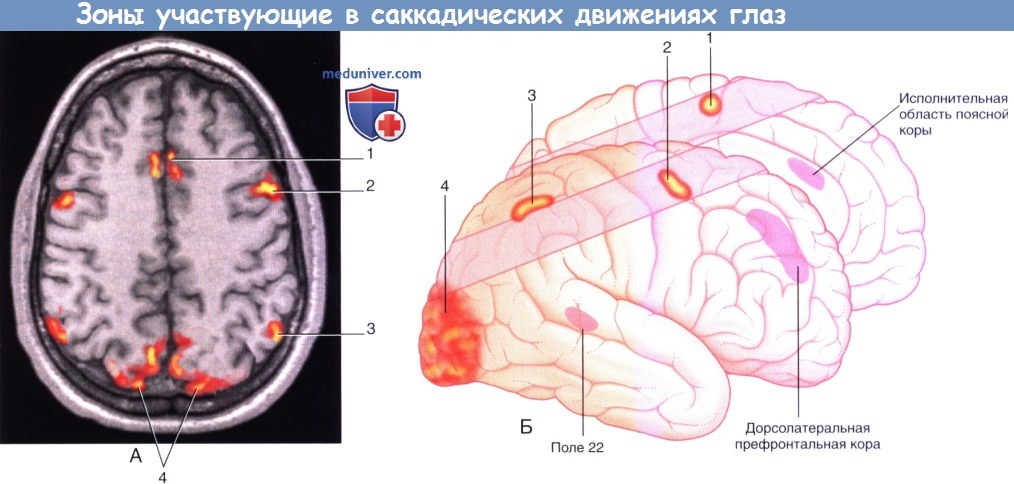 Зоны коры больших полушарий, участвующие в саккадических движениях глаз.
Зоны коры больших полушарий, участвующие в саккадических движениях глаз.
(1) Дополнительное глазодвигательное поле.
(2) Фронтальное глазодвигательное поле.
(3) Теменное глазодвигательное поле.
(4) Ассоциативная зрительная кора.
в) Синдром ригидного человека (СРЧ). Редкое, но легко распознаваемое заболевание, известное как синдром ригидного человека (СРЧ), или синдром мышечной скованности, — аутоиммунное заболевание центральной нервной системы, связанное с наличием циркулирующих антител к глутаматдекарбоксилазе (GAD65),— ключевому ферменту, преобразующему глутамат в ГАМК. Синдром ригидного человека (СРЧ) проявляется скованностью мышц с эпизодическими мышечными спазмами (вызванными одновременным сокращением мышц синергистов и антагонистов, преимущественно в проксимальных отделах конечностей и осевой мускулатуре) и боязнью выполнения определенных задач. В норме контроль верхних моторных нейронов происходит за счет тонической активности прилежащих тормозных ГАМК-ергических вставочных нейронов.
Одни зоны коры поражаются сильнее других, а клинические проявления связаны с нарушением функции этих ГАМК-ергических нейронов, что приводит к гипервозбудимости коры. В настоящее время уточняют роль циркулирующих антител в патогенезе синдрома ригидного человека (СРЧ).
г) Резюме. Кора больших полушарий имеет одновременно пластинчатую и колончатую организацию. К двум основным типам клеток относят пирамидные и непирамидные клетки (вставочные нейроны). Пирамидные клетки расположены в слоях (пластинках) II, III, а также V (в виде веретеновидных клеток) и VI. Слой IV богат шиповатыми звездчатыми клетками (модифицированными пирамидными нейронами). Мелкие пирамидные клетки связывают извилины одного полушария, средние пирамидные клетки связывают между собой соответствующие зоны двух полушарий, а от крупных пирамидных клеток отходят волокна к таламусу, стволу мозга и спинному мозгу. Все корковые волокна считают возбуждающими; шиповатые звездчатые клетки также считают возбуждающими пирамидными клетками.
Корковые вставочные нейроны—тормозные. Колончатая организация представлена колоннами клеток, которые рассматривают в качестве первичной функциональной единицы обработки информации в коре; они состоят из определенных нейронных «микроцепей».
Части тела в соматосенсорной коре представлены инвертированными. Важная входящая информация поступает от вентрального заднего ядра таламуса, а направляется к первичной моторной коре и нижней теменной коре. В первичную зрительную кору входит коленчато-шпорный путь. Клеточные реакции различной сложности зависят от передачи информации с более простых типов клеток на более сложные. Свойство ассоциативных зрительных зон—распознавание черт, например цвета, формы и движения. Обработка цвета и формы продолжается в коре нижележащей височной доли, а движений—в задней теменной доле. Первичная слуховая кора занимает верхнюю поверхность верхней височной извилины, а ассоциативная слуховая кора расположена с латеральной стороны от нее.
Первичная моторная кора занимает прецентральную извилину. Она дает начало большей части волокон пирамидного тракта, а части тела в ней представлены инвертированными. Основная информация подходит к ней от соматосенсорной коры, мозжечка (через вентральное заднее ядро таламуса), премоторной и дополнительной моторной зон. Премоторная кора отвечает преимущественно на внешние раздражители, а дополнительная моторная зона — на внутренние стимулы. Дорсолатеральная префронтальная кора контролирует четыре различных корковых зоны, в различной степени участвующих в создании противоположных саккад.
Редактор: Искандер Милевски. Дата публикации: 22.11.2018
Источник