За эмоции отвечают голубое пятно покрышки среднего мозга черная субстанция среднего мозга
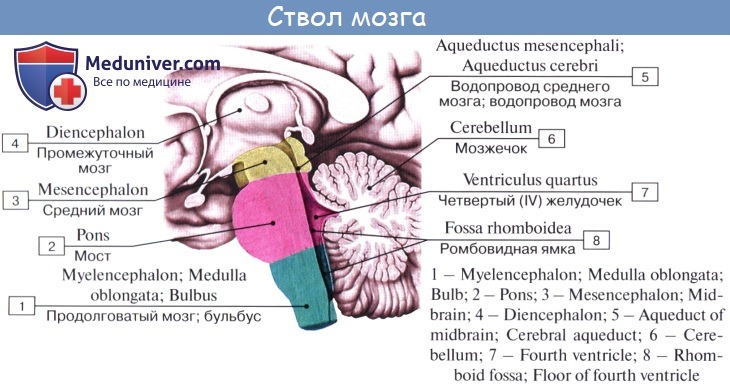
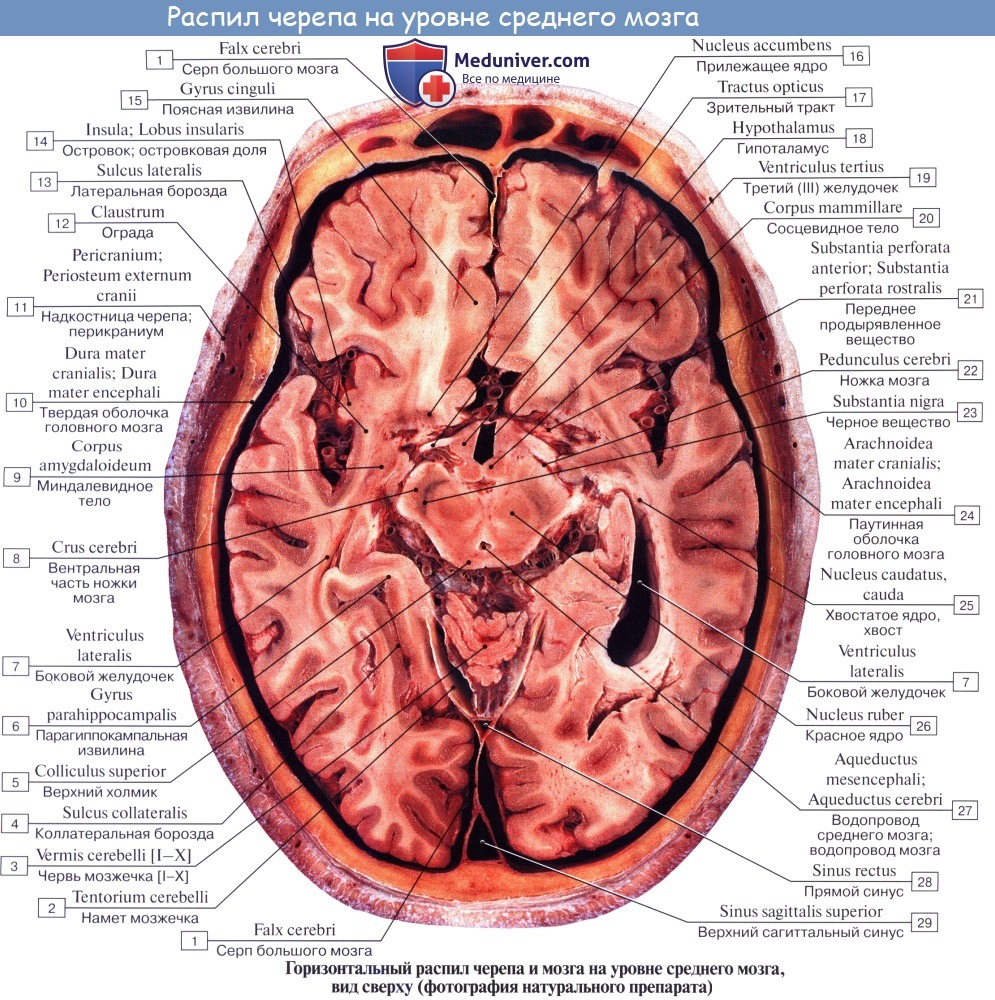
Средний мозг, mesencephalon, развивается в процессе филогенеза под преимущественным влиянием зрительного рецептора, поэтому важнейшие его образования имеют отношение к иннервации глаза. Здесь же образовались центры слуха, которые вместе с центрами зрения в дальнейшем разрослись в виде четырех холмиков крыши среднего мозга.
С появлением у высших животных и человека коркового конца слухового и зрительного анализаторов в коре переднего мозга слуховые и зрительные центры среднего мозга сами попали в подчиненное положение и стали промежуточными, подкорковыми. С развитием у высших млекопитающих и человека переднего мозга через средний мозг стали проходить проводящие пути, связывающие кору конечного мозга со спинным (ножки мозга).
В результате в среднем мозге человека имеются:
1) подкорковые центры зрения и ядра нервов, иннервирующих мышцы глаза;
2) подкорковые слуховые центры;
3) все восходящие и нисходящие проводящие пути, связывающие кору головного мозга со спинным и идущие транзитно через средний мозг;
4) пучки белого вещества, связывающие средний мозг с другими отделами центральной нервной системы.
Соответственно этому средний мозг, являющийся у человека наименьшим и наиболее просто устроенным отделом головного мозга, имеет две основные части: крышу, где располагаются подкорковые центры слуха и зрения, и ножки мозга, где преимущественно проходят проводящие пути.
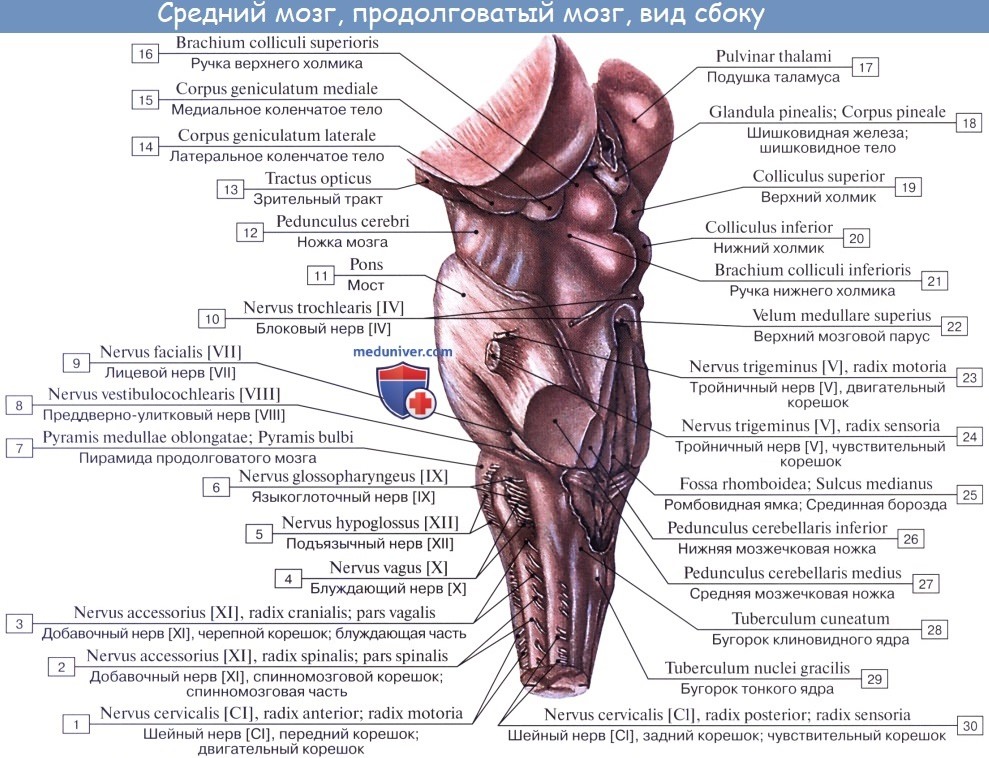
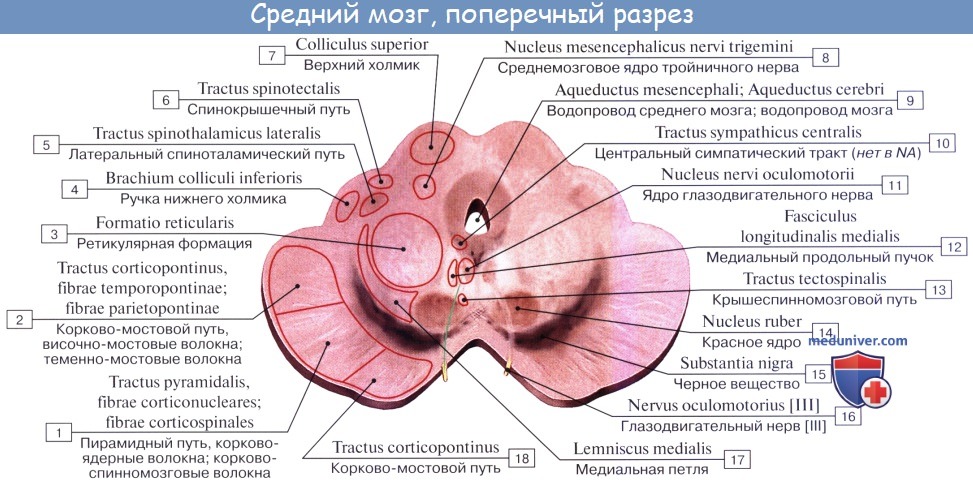
Дорсальная часть, крыша среднего мозга, tectum mesencephali.
Она скрыта под задним концом мозолистого тела и подразделяется посредством двух идущих крест-накрест канавок — продольной и поперечной — на четыре холмика, располагающихся попарно.
Верхние два холмика, colliculi superiores, являются подкорковыми центрами зрения, оба нижних, colliculi inferiores,— подкорковыми центрами слуха. В плоской канавке между верхними бугорками лежит шишковидное тело. Каждый холмик переходит в так называемую ручку холмика, brachium colliculi, направляющуюся латерально, кпереди и кверху, к промежуточному мозгу. Ручка верхнего холмика, brachium colliculi superioris, идет под подушкой, pulvinar, таламуса к латеральному коленчатому телу, corpus geniculatum laterale.
Ручка нижнего холмика, brachium colliculi inferioris, проходя вдоль верхнего края trigonum lemnisci до sulcus lateralis mesencephali, исчезает под медиальным коленчатым телом, corpus geniculatum mediale. Названные коленчатые тела относятся уже к промежуточному мозгу.
Вентральная часть, ножки мозга, pedunculi cerebri, содержит все проводящие пути к переднему мозгу.
Ножки мозга имеют вид двух толстых полуцилиндрических белых тяжей, которые расходятся от края моста под углом и погружаются в толщу полушарий большого мозга.
Полость среднего мозга, являющаяся остатком первичной полости среднего мозгового пузыря, имеет вид узкого канала и называется водопроводом мозга, aqueductus cerebri. Он представляет узкий, выстланный эпендимой канал 1,5 — 2,0 см длиной, соединяющий IV желудочек с III. Дорсально водопровод ограничивается крышей среднего мозга, вентрально — покрышкой ножек мозга.
На поперечном разрезе среднего мозга различают три основные части:
1) пластинку крыши, lamina tecti;
2) покрышку, tegmentum, представляющую верхний отдел pedunculi cerebri;
3) вентральный отдел pedunculi cerebri, или основание ножки мозга, basis pedunculi cerebralis.
Соответственно развитию среднего мозга под влиянием зрительного рецептора в нем заложены различные ядра, имеющие отношение к иннервации глаза.
У низших позвоночных верхнее двухолмие служит главным местом окончания зрительного нерва и является главным зрительным центром. У млекопитающих и у человека с переносом зрительных центров в передний мозг остающаяся связь зрительного нерва с верхним холмиком имеет значение только для рефлексов. В ядре нижнего холмика, а также в медиальном коленчатом теле оканчиваются волокна слуховой петли (lemniscus lateralis). Крыша среднего мозга имеет двустороннюю связь со спинным мозгом — tractus spinotectalis и tractus tectobulbaris et tectospinalis. Последние после перекреста в покрышке идут к мышечным ядрам в продолговатом и спинном мозге. Это так называемый зрительно-звуковой рефлекторный путь, о котором говорилось при описании спинного мозга. Таким образом, пластинку крыши среднего мозга можно рассматривать как рефлекторный центр для различного рода движений, возникающих главным образом под влиянием зрительных и слуховых раздражений.
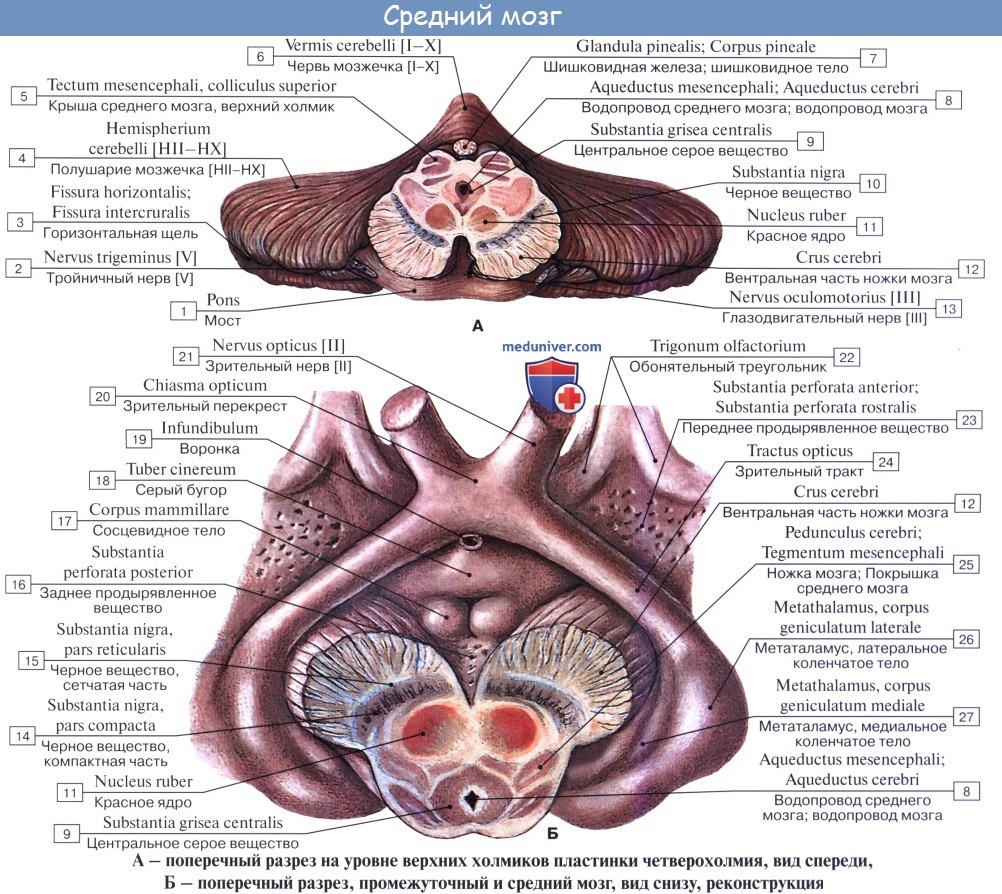
Водопровод мозга окружен центральным серым веществом, имеющим по своей функции отношение к вегетативной системе. В нем, под вентральной стенкой водопровода, в покрышке ножки мозга заложены ядра двух двигательных черепных нервов — n. oculomotorius (III пара) на уровне верхнего двухолмия и n. trochlearis (IV пара) на уровне нижнего двухолмия. Ядро глазодвигательного нерва состоит из нескольких отделов соответственно иннервации нескольких мышц глазного яблока.
Медиально и кзади от него помещается еще небольшое, тоже парное, вегетативное добавочное ядро, nucleus accessories, и непарное срединное ядро. Добавочное ядро и непарное срединное ядро иннервируют непроизвольные мышцы глаза, m. ciliaris и m. sphincter pupillae. Эта часть глазодвигательного нерва относится к парасимпатической системе. Выше (ростральнее) ядра глазодвигательного нерва в покрышке ножки мозга располагается ядро медиального продольного пучка.
Латерально от водопровода мозга находится ядро среднемозгового тракта тройничного нерва, nucleus mesencephalicus n. trigemini.
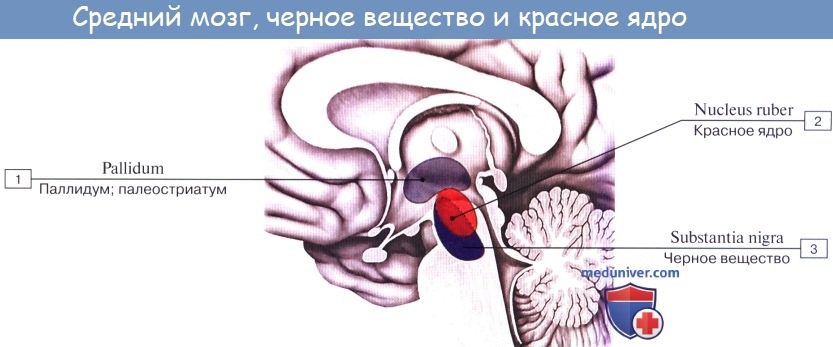
Ножки мозга делятся, как уже отмечалось, на вентральную часть, или основание ножки мозга, basis pedunculi cerebralis, и покрышку, tegmentum. Границей между ними служит черное вещество, substantia nigra, обязанное своим цветом содержащемуся в составляющих его нервных клетках черному пигменту — меланину.
Покрышка среднего мозга, tegmentum mesencephali, — часть среднего мозга, расположенная между его крышей и черным веществом (substantia nigra) ножек мозга.
От нее отходит tractus tegmentalis centralis — центральный покрышечный путь — проекционный нисходящий нервный путь, расположенный в центральной части покрышки среднего мозга. Он содержит волокна, идущие от таламуса, бледного шара, красного ядра и ретикулярной формации среднего мозга к ретикулярной формации и оливе продолговатого мозга; относится к экстрапирамидной системе.
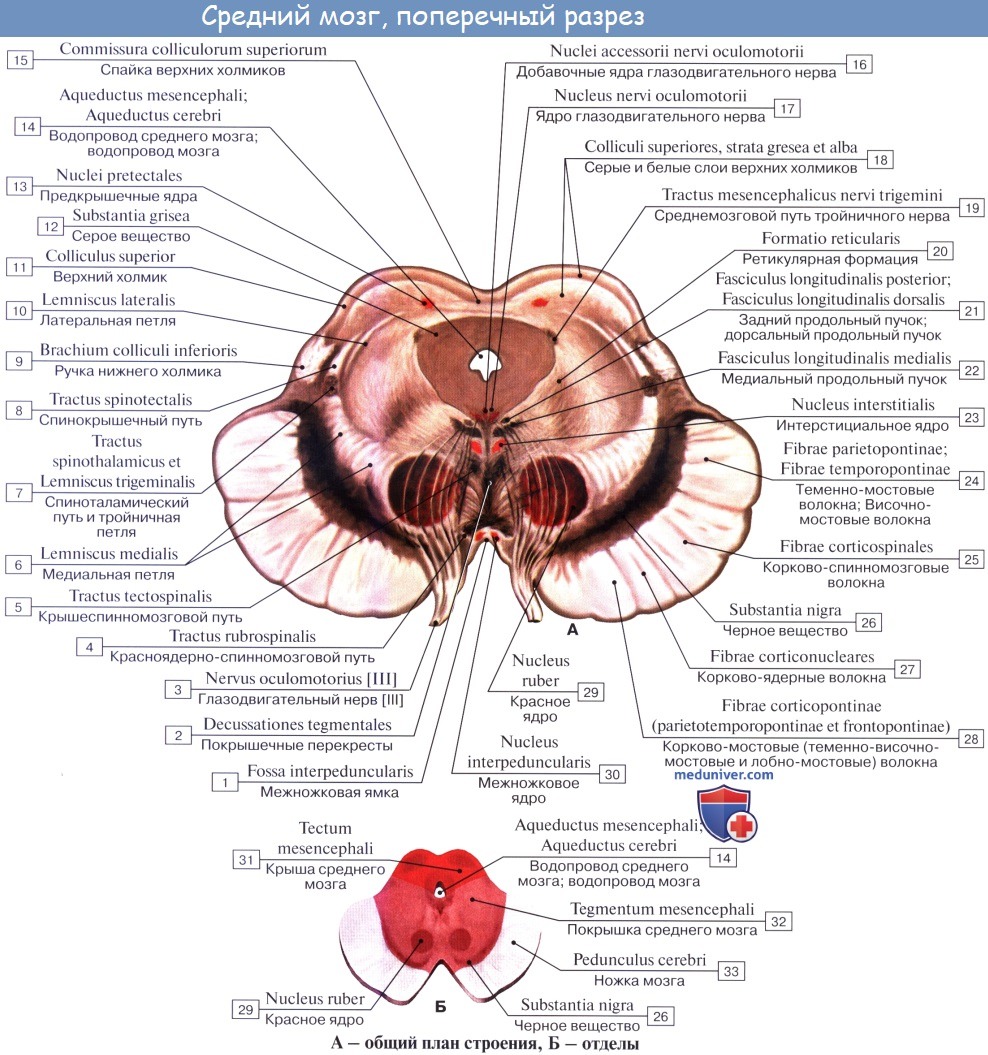
Substantia nigra простирается на всем протяжении ножки мозга от моста до промежуточного мозга; по своей функции относится к экстрапирамидной системе.
Расположенное вентрально от substantia nigra основание ножки мозга содержит продольные нервные волокна, спускающиеся от коры полушария большого мозга ко всем нижележащим отделам центральной нервной системы (tractus corticopontmus, corticonuclearis, corticospinalis и lдр.).
Tegmentum, находящаяся дорсально от substantia nigra, содержит преимущественно восходящие волокна, в том числе медиальную и латеральную петли. В составе этих петель восходят к большому мозгу все чувствительные пути, за исключением зрительного и обонятельного.
Среди ядер серого вещества самое значительное — красное ядро, nucleus ruber. Это удлиненное колбасовидное образование простирается в покрышке ножки мозга от гипоталамуса промежуточного мозга до нижнего двухолмия, где от него начинается важный нисходящий тракт, tractus rubrospinal, соединяющий красное ядро с передними рогами спинного мозга. Пучок этот после выхода из красного ядра перекрещивается с аналогичным пучком противоположной стороны в вентральной части срединного шва — вентральный перекрест покрышки.
Nucleus ruber является весьма важным координационным центром экстрапирамидной системы, связанным с остальными ее частями. К нему проходят волокна от мозжечка в составе верхних ножек последнего после их перекреста под крышей среднего мозга, вентрально от aqueductus cerebri, а также от pallidum — самого нижнего и самого древнего из подкорковых узлов головного мозга, входящих в состав экстрапирамидной системы. Благодаря этим связям мозжечок и экстрапирамидная система через посредство красного ядра и отходящего от него tractus rubrospinal оказывают влияние на всю скелетную мускулатуру в смысле регуляции бессознательных автоматических движений.
В покрышку среднего мозга продолжаются также ретикулярная формация, formatio reticularis, и fasciculus longitudindlis medialis. Последний берет начало в различных местах. Одна из его частей начинается из вестибулярных ядер, проходит на той и другой стороне по бокам средней линии, непосредственно под серым веществом дна водопровода и IV желудочка, и состоит из восходящих и нисходящих волокон, идущих к ядрам III, IV, VI и XI черепных нервов.
Медиальный продольный пучок является важным ассоциативным путем, связующим различные ядра нервов глазных мышц между собой, чем обусловливаются сочетанные движения глаз при отклонении их в ту или другую сторону. Функция его связана также с движениями глаз и головы, возникающими при раздражении аппарата равновесия.
Видео анатомия среднего мозга от А.А. Стрелкова
— Вернуться в оглавление раздела «Анатомия нервной системы.»
Источник
За эмоции отвечают голубое пятно покрышки среднего мозга черная субстанция среднего мозга
В 60—70-х годах с помощью гистохимических и радиоиммунологических методов было установлено, что в мозге имеются дифференцированные нейрохимические системы — норадренергическая, дофаминергическая, серотонинергическая и др. Они представляют собой комплекс мозговых структур, функция которых определяется наличием общего нейротрансмиттера и рецепторов, взаимодействующих с ним. В одних из структур, входящих в нейрохимическую систему, расположены тела нейронов, в других оканчиваются терминали нервных клеток. В последнем случае говорят о проекциях на те или иные мозговые образования. В одних и тех же структурах мозга могут располагаться клетки и проекции нескольких нейрохимических систем.
Дофаминергическая система. В этой системе мозга различают 7 отдельных подсистем (систем, трактов): нигростриатную, мезокортикальную, мезолимбическую, тубероинфундибулярную, инцертогипоталамическую, диенцефалоспинальную и ретинальную. Из них первые 3 являются основными (рис. 8). Тела нейронов нигростриатной, мезокортикальной и мезолимбической систем расположены на уровне среднего мозга, образуют комплекс нейронов черной субстанции и вентрального поля покрышки. Они составляют непрерывную клеточную сеть, проекции которой частично перекрываются, поскольку аксоны этих нейронов идут вначале в составе одного крупного тракта (медиального пучка переднего мозга), а оттуда расходятся в разные мозговые структуры. Формирование нигростриатной, мезолимбической и мезокортикальной систем определяется областями, где оканчиваются аксоны дофаминергических нейронов, т.е. локализацией их проекций. Некоторые авторы объединяют мезокортикальную и мезолимбическую подсистемы в единую систему. Более обоснованным является выделение мезокортикальной и мезолимбической подсистем соответственно проекциям в лобную кору и лимбические структуры мозга (см. рис. 8).

Нигростриатная система. Нигростриатный тракт является самым мощным в дофаминергической системе мозга. Аксонами нейронов этого тракта выделяется около 80 % мозгового дофамина. Тела дофаминовых нейронов, образующих этот путь, находятся в основном в компактной части черной субстанции, но часть волокон берет начало также от нейронов латерального отдела вентрального поля покрышки среднего мозга.
Клетки компактной части черной субстанции дают проекции в дорсальный стриатум (полосатое тело), а клетки вентрального поля покрышки — в вентральный стриатум. Наиболее плотно расположены дофаминергические волокна в стриатуме — они начинаются от латеральных отделов черной субстанции того же полушария. Эти волокна оканчиваются на нейронах хвостатого ядра и скорлупы, т.е. в неостриатуме. Дофаминергическую иннервацию получают также другие структуры, в частности базальные ганглии — бледный шар (палеостриатум) и субталамическое ядро. У хвостатого ядра более плотная иннервация отмечается в головке и значительно меньше плотность дофаминергических проекций в каудальной части.

Мезокортикальная система. Тела нейронов, образующих мезокортикальный тракт, находятся в вентральной части покрышки среднего мозга, а основные проекции этих нейронов достигают лобной (преимущественно префронтальной, поле 10 по Бродману — рис. 9) коры. Соответствующие окончания расположены в основном в глубоких слоях лобной коры ( V — VI ). Мезокортикальная дофаминовая система оказывает большое влияние на активность нейронов, образующих корково-корковые, корково-таламичес-кие и корково-стриатные пути.
Мезолимбическая система. Источники дофаминергических проекций, т.е. тела нейронов этой системы, расположены в вентральном поле покрышки среднего мозга и частично в компактной части черной субстанции. Их отростки идут в поясную извилину, энториальную кору, миндалину, обонятельный бугорок, аккумбентное ядро, гиппокамп, парагиппокампальную извилину, перегородку и другие структуры лимбической системы мозга. Имея обширные связи, Мезолимбическая система опосредовано проецируется также на лобную кору и гипоталамус. Это определяет широкие функции мезолимбической системы, которая участвует в механизмах памяти, эмоций, обучения и нейроэндокринной регуляции.
Другие тракты. Тубероинфундибулярный тракт образован аксонами нейронов, расположенных в аркуатном ядре гипоталамуса. Отростки таких нейронов достигают наружного слоя срединного возвышения. Этот тракт осуществляет контроль секреции пролактина. Дофамин тормозит его секрецию и поэтому содержание пролактина в плазме крови служит косвенным показателем функции дофаминергической системы мозга, что часто используют для оценки влияния на нее психофармакологических средств. Инцертогипоталамический тракт начинается от zona incerta и оканчивается в дорсальном и переднем отделах медиального таламуса, а также в перивентрикулярной области. Он принимает участие в нейроэндокринной регуляции. Источником проекций диенцефалоспинального тракта являются нейроны заднего гипоталамуса, отростки которых достигают задних рогов спинного мозга. Ретинальный тракт расположен в пределах сетчатки глаза. Особенности этого тракта делают его среди других дофаминергических трактов достаточно автономным.
Приведенная дифференциация дофаминергических образований мозга не является абсолютной, так как проекции дофаминергических нейронов разных трактов «перекрываются». Кроме того, в мозге отмечается и диффузное распределение дофаминергических элементов (отдельных клеток с отростками).
Дофаминергические системы мозга созревают преимущественно в постанальном периоде.
Дофаминовые терминали образуют синапсы преимущественно на шипиках и стволах дендритов — это аксошипиковые и аксодендритные синапсы (их более 90 %). Лишь единичные синапсы (менее 10 %) расположены на телах нейронов (аксосоматические) и на аксонах (аксо-аксональные).
Основными типами дофаминовых рецепторов являются Д1- и Д2-ре-цепторы. Недавно были открыты также рецепторы ДЗ, Д4 и Д5. Они все находятся главным образом на постсинаптической мембране. Но в дофаминергической системе существуют также ауторецепторы, расположенные на теле нейронов, аксонах, дендритах и терминалях, которые реагируют на собственный дофамин, регулируя его синтез и выделение. Их стимуляция приводит к снижению активности дофаминовых нейронов.
Большая часть охарактеризованных дофаминовых рецепторов относится к Д2-рецепторам. О функции Д1-рецепторов известно меньше. Их от Д2-рецепторов отличает способность стимулировать активность фермента аденилатциклазы, которая в свою очередь участвует в синтезе второго мессенджера — цАМФ. Д1- и Д2-рецепторы существуют в двух формах — высоко- и низкоаффинной, что определяется по их способности к связыванию агонистов и антагонистов. Было показано, что Д2-рецепторы сродство к бутирофенонам, в то время как Д1-рецепторы такой способностью не обладают.
Д2-рецепторы преобладают в стриатуме — хвостатом ядре и скорлупе, но имеются также в поясной извилине и коре островка. В стриатуме Д2 обнаружены не только на дофаминергических, но и на холинергических нейронах. Это объясняет сопряженное выделение ацетилхолина при введении агонистов дофамина. Картирование Д1-рецепторов дало менее убедительные результаты, однако было установлено, что они преобладают в коре больших полушарий, особенно в области префронтальной коры, где имеются и Д2-рецепторы. Д1-рецепторы есть и в стриатуме.
Рецепторы ДЗ, Д4, Д5 были открыты относительно недавно. Структурные особенности и фармакологические свойства рецепторов ДЗ близки к таковым рецепторов Д2. Рецептор Д4 также имеет сходство с Д2 и ДЗ, а рецептор Д5 — с Д1.
Различные психофармакологические препараты, будучи агонистами или антагонистами, относительно избирательно влияют на Д1- и Д2-рецепторы. Так, фенотиазины блокируют Д1- и Д2-рецепторы; галоперидол и пимозид являются более сильными блокаторами этих рецепторов; сильным антагонистом считается также сульпирид. В противоположность им флупентиксол высокоаффинен к Д1-рецепторам.
Большое значение для понимания механизмов действия нейролептиков на уровне дофаминовых рецепторов придают их молекулярному строению. Оказалось, что молекулы дофамина и фенотиазина, по данным рентгеноструктурного анализа, обладают определенным структурным сходством. Есть также сходство в молекулах дофамина и амфетамина (агонист дофамина). Поэтому нейролептики способны устранять психотомиметические эффекты последнего.
Длительное введение нейролептиков приводит к повышению связывания дофаминовых рецепторов и усилению поведенческих реакций на агонисты дофамина. Это явление трактуется как гиперчувствительность дофаминовых рецепторов. С ним связывают развитие побочных явлений при лечении нейролептиками, в частности с гиперчувствительностью рецепторов в стриатуме — развитие экстрапирамидных расстройств. Но открытие атипичных нейролептиков внесло некоторые коррективы в ранее существовавшие представления. Оказалось, что такой атипичный нейролептик, как сульпирид, являясь высокоспецифичным по отношению к Д2-рецепторам (в отличие от Хлорпромазина, галоперидола и др.), действуя на клетки вентральной области покрышки, иннервирующие лимбическую кору и кору больших полушарий, не влияет на нейроны черной субстанции, проецирующиеся на полосатое тело. Этим объясняется редкость возникновения лекарственного паркинсонизма при лечении сульпиридом.
Такой атипичный нейролептик, как клозапин (лепонекс), связывает не только Д2-, но в большей степени Д1-рецепторы и рецепторы других нейрохимических систем (серотониновые, ацетилхолиновые и др.).
Серотонинергическая система. Нейроны, являющиеся источником путей серотонинергической системы, находятся главным образом в переднем (ростральном) и заднем (каудальном) ядрах шва мозгового ствола. Они образуют группы клеток, расположенные от передней части мезенцефалона до нижних отделов продолговатого мозга. Отростки этих клеток широко разветвлены и проецируются на большие области коры переднего мозга, его желудочковую поверхность, мозжечок, спинной мозг и образования лимбической системы (рис. 10).

В ядрах шва серотонинергические нейроны локализуются вместе с нейронами другой химической принадлежности (ГАМКергическими, выделяющими субстанцию P, энкефалиновыми и др.).
В основе функционирования серотонинергической системы лежит выделение серотонина, или 5-окситриптамина (5- hydroxytriptamine , 5- HT ) в синаптическую щель. В последней он частично инактивируется и частично захватывается обратно пресинаптической терминалью. Именно на эти процессы влияют антидепрессанты последней генерации, которые получили название ингибиторов обратного захвата серотонина.
В серотонинергической системе имеется 2 типа рецепторов — 5-НТ1 и 5-НТ2. Существует тенденция разделять их на подтипы: 5-НТ1А и т.д. Один из относительно новых анксиолитиков — буспирон — способен стимулировать именно 5-НТ1А-рецепторы.
С нарушением функции серотонинергической системы связывают развитие психических нарушений, проявляющихся депрессией и тревогой.
Норадренергическая система. Источником норадренергических путей в мозге являются группы клеток, расположенных в мозговом стволе и ретикулярной формации. Они включают клетки голубого пятна ( locus ceruleus ), вентромедиальной части покрышки и др. Отростки таких клеток сильно разветвлены и коллатеризированы. Области, на которые распространяются восходящие проекции этих клеток, захватывают ствол мозга, гипоталамус, таламус и разные отделы коры, а нисходящие достигают спинного мозга (рис. 11). Восходящие норадренергические проекции являются компонентом восходящих активирующих систем.

Адренергические рецепторы разделяются на — и ß 1 -, а последние на ß 1 и ß 2 . Рецепторы ß 1 локализуются на нейроне, а ß 2 — на клетках глии и сосудов. Агонистом ß 1 -рецепторов является норадреналин, а ß 2 -рецепторы более чувствительны к адреналину.
Рецепторы типов 1 и 2 хорошо изучены фармакологически. Специфические ингибиторы 1 -рецепторов обладают антигипертензивными свойствами, 2 -рецепторы в большой степени определяют активность центральной и периферической адренергической систем. Пресинаптические а 2 -рецепторы на норадренергических терминалях тормозят выделение норадреналина, имея отношение и к регуляции кровяного давления. Об этом свидетельствует, в частности, влияние клонидина, который будучи антигипертензивным средством уменьшает также симптомы абстиненции при алкоголизме и наркоманиях.
Холинергическая система. Эта система состоит из нейронов, выделяющих ацетилхолин — ее нейротрансмиттер. Холинергические нейроны достаточно широко представлены в мозге, но центральными областями ее являются кора (лобная, теменная, височная), гиппокамп, хвостатое тело и ядро Мейнерта (базальное ядро Мейнерта), функции которых имеют отношение к когнитивным процессам, включая память.
Функционирование холинергической системы определяют мускариновые ацетилхолиновые рецепторы — Ml и М2, различающиеся по вторичным внутриклеточным процессам (вторичные мессенджерные системы). Последнее делает их сходными с гистаминовыми рецепторами.
ГАМКергическая система. ГАМК (гамма-аминомасляная кислота) синтезируется из глютаминовой кислоты при участии декарбоксилазы. Этот фермент локализуется в нейронах, использующих ГАМК как тормозящий нейротрансмиттер. К ГАМКергической системе относятся интернейроны коры, афферентные волокна, идущие от полосатого тела к бледному шару и черной субстанции, а также клетки Пуркинье мозжечка. С ГАМКергической системой связано и тормозящее влияние глицина, локализация которого ограничена нейронами ствола мозга и спинного мозга. Быстрое развитие торможения нейрональной активности путем активации глициновых и ГАМКергических рецепторов опосредовано открытием ионных хлорных каналов, что позволяет ионам С1 проникать в нейрон, вызывая их гиперполяризацию. В результате этого они становятся менее чувствительными к стимулам. С функцией ГАМК-рецепторов связано действие бензодиазепинов — их агонистов.
ГАМК-рецепторы также имеют подтипы. Так, с бензодиазепинами реагирует ГАМК В -рецептор, агонист его баклофен является сильным антиспастическим агентом.
Структуры мозга, в которых обнаруживается наибольшее содержание ГАМК, имеют и высокий уровень дофамина. Поэтому во многих предположениях об участии ГАМК в патогенезе психических расстройств этот нейротрансмиттер рассматривается в связи с изменением функции дофаминергических нейронов. Существует мнение, что ГАМК не только нейротрансмиттер, но и синаптический модулятор на уровне дофаминового рецептора. С функцией ГАМКергической системы связывают также положительное влияние глицина в некоторых случаях шизофрении (в частности, при резистентности к нейролептикам).
Помимо структурно организованных нейрохимических систем, большую роль в функциях мозга играют и другие нейротрансмиттеры и рецепторы, в числе которых должны быть названы возбуждающие аминокислоты и гистамин.
Возбуждающие аминокислоты. Функция этих аминокислот имеет отношение к деятельности перечисленных нейрохимических систем, особенно ГАМКергической. К возбуждающим аминокислотам относятся глютамат и аспартат, которые рассматриваются как основные возбуждающие нейротрансмиттеры в мозге. Опосредуют эффекты глютамата три главных рецептора, дифференцирующихся по их основным специфическим агонистам, — N -метил-О-аспартат ( NMDA ), квискуалат и каинат, т.е. имеются NMDA -, квисквалатные и каинатные рецепторы. С функцией этих рецепторов, особенно NMDA и каинатных, связывают действие некоторых антисудорожных средств и психотомиметиков (например, фенциклидина).
Продолжительная стимуляция таких рецепторов может вызывать дегенерацию нейронов, в то время как аксоны тех же клеток, особенно на отдаленных от тела нейрона участках, остаются сохранными. Такая реакция получила название «эксайтотоксичности» ( exitotoxicity ). Она может быть вызвана, например, инъекцией в полосатое тело каиновой или хинолиновой кислоты (эндогенный метаболит триптофана), которые действуют и на NMDA -рецепторы, приводя к развитию нейродегенеративного процесса, напоминающего гистологическую картину хореи Гентингтона.
Гистамин. Это вещество содержится в тканях и биологических жидкостях организма. Оно имеется и в тканях мозга, причем его содержание, например, в гипофизарной области превосходит уровень других биологических аминов. Большое количество гистамина в гипоталамусе, сосудистом сплетении, эпифизе и меньшее — в таламусе, среднем мозге и коре.
До последнего времени оставались сомнения в отношении принадлежности гистамина к нейротрансмиттерам. Но изучение его рецепторов и их фармакологических свойств, особенно с помощью новейших методов при жизненной визуализации мозговых структур и связывания рецепторов соответствующими агонистами, позволило рассматривать это физиологически активное вещество как нейротрансмиттер, которому соответствуют специфические рецепторы.
Различают два типа гистаминовых рецепторов — HI и Н2. Они по-разному влияют на внутриклеточные вторично-мессенджерные процессы: Н1-рецепторы стимулируют фосфоинозитидиновую систему, а Н2-рецепторы активируют аденилатциклазную систему. Рецепторы Н2 имеются в коре и гиппокампе. Они блокируются трициклическими антидепрессантами. С Н1-рецепторами связывают действие антигистаминовых препаратов, которые, являясь их антагонистами, блокируют их. Блокаторы Н2 используют для подавления желудочной секреции, но при их длительном применении возможно развитие органического мозгового синдрома (особенно у пожилых женщин).
Нейропептиды. В течение последних двух десятилетий большое внимание исследователей привлекают нейропептиды, которые служат основой межклеточных взаимодействий различной модальности, так как выступают в качестве нейрогормонов, нейротрансмиттеров, нейромодуляторов и, возможно, как химические переносчики специфической информации между нервными клетками.
Для психиатрии особый интерес представляют нейропептиды, составляющие группу эндогенных опиоидов, — эндогенных веществ, обладающих свойствами морфина. Они получили название эндорфинов и энкефалинов. К ним относятся а-, p- и у- эндорфины, метионин-эндорфин и лейкин-эндорфин. По структуре эндорфины и энкефалины представляют собой различной величины фрагменты молекулы липотропина.
Эндогенные опиаты взаимодействуют со специфическими рецепторами к ним. Различают три типа таких рецепторов — дельта, мю и каппа ( , , ). Действие экзогенных или эндогенных лигандов с этими рецепторами вызывает соответствующие физиологические эффекты. Так, активация -рецептора вызывает аналгезию, агонисты -рецепторов приводят к возникновению галлюцинаций и дисфории. Молекула -эндорфина является смешанным лигандом, реагирует с — и -рецепторами.
Опиоидные пептиды очень широко представлены в мозге — в телах нервных клеток, отростках и терминалях. Они были обнаружены в височной и энториальной коре, в гипоталамусе, мозолистом теле, перивентрикулярной части таламуса, перегородке, полосатом теле, хвостатом ядре, обонятельном бугорке и других структурах. Области, в которых распределены разные эндогенные опиаты, имеющие и разные типы рецепторов, «перекрываются» между собой, а также с зонами локализации структурно-химических компонентов других систем, в частности дофаминовой. На основе имеющихся нейрохимических и фармакологических данных о тесной структурной и функциональной связи дофаминергической и эндорфиновой систем мозга появились предположения о модулирующей роли эндорфинов в процессе передачи информации через дофаминовые синапсы на пре- и постсинаптическом уровне.
К другим нейропептидам относятся вещества, обладающие нейрогормональной функцией — вазопрессин, окситоцин, нейротензин, холецистокинин, соматостатин, тиреотропный гормон и др. Нейрогормональной функцией обладают и нейропептиды, вырабатываемые нейронами гипоталамуса, так называемые рилизинг-гормоны, стимулирующие высвобождение тропных гормонов передней доли гипофиза. Некоторые из них существенно влияют на функцию мозга. Например рилизинг-гормон тиреотропина улучшает состояние больных депрессией, а соматостатин (ингибитор выброса гормона роста), напротив, способен усугублять депрессию.
Нейротрансмиттерная функция нейропептидов изучена меньше. Осуществление этой функции связывают с веществом P. Это вещество изменяет мембранные потенциалы нейрона, ослабляет аналгезию, вызванную морфином, и абстиненцию при морфинной наркомании.
Открытие С. Pert , S . Snyder (1973) эндогенных морфинов и опиатных рецепторов по своему значению вышло далеко за пределы проблемы нейропептидов, поскольку оно положило начало изучению эндогенных соединений с выраженным психотропным эффектом (некоторые эндорфины активнее морфина), которые имеют специфические рецепторные структуры к ним. Большое внимание в связи с этим привлекли работы Н. M
Из вышеизложенного видно, что существует тесная связь в функции отдельных нейрохимических систем, особенно в виде модуляции на уровне действия нейротрансмиттеров.
Источник