Смешанные чувства
Вариант восприятия (но не ви́дения) буквенно-цифрового набора синестетом: символы он видит в «правильном» цвете — в котором они изображены на самом деле, — но одновременно воспринимает и свои собственные цвета, рождающиеся в результате активации «неправильных» зон мозга за компанию с «правильными».
Автор
Редакторы
Восприятие окружающего мира формируется из различных чувств: зрение, слух, обоняние, осязание и вкус. Каждый человек может разделить потоки информации, которые приходят от разных органов чувств: мы не путаем зрительный образ котлеты и запах, идущий от нее. Однако существует феномен, при котором потоки информации смешиваются, звуки обретают цвет, а тактильные ощущения вызывают эмоции. Он называется синестезией.
«Темно-коричневое отполированное Я»
Синестезия — это явление, при котором сенсорный или когнитивный стимул вызывает ответ не только в соответствующей сфере, но и непроизвольное и неконтролируемое появление сенсорного сигнала другой модальности (другой сферы). Человек слышит какой-то звук, например, ноту, взятую на пианино, и у него возникает ощущение синего цвета.
Тут сразу нужно сделать несколько оговорок. Во-первых, синестеты — люди, обладающие способностью к синестезии, — здоровы. Эта особенность их нервной системы не требует никакого вмешательства врачей. Во-вторых, тип синестезии, например, связь между буквой и цветом или между текстурой ткани, которой касается синестет, и эмоциональным ощущением, возникающим при этом, устойчивы. Как правило, они не меняются на протяжении всей жизни (хотя возможны исключения). Это — главное качественное отличие синестезии от ассоциаций. Ассоциации зависят от личного опыта человека и могут меняться, а синестетические ощущения стабильны и не связаны с тем, что он переживал ранее. В-третьих, оставаясь довольно умозрительным феноменом, синестезия была достаточно хорошо изучена. Известно, что у женщин она встречается чаще, чем у мужчин.
Считается, что первое научное описание синестезии принадлежит Френсису Гальтону: оно было изложено в виде краткой заметки в 1892 году. Хотя существует работа Джорджа Тобиаса Людвига Сакса, опубликованная в 1812 году, где тот описывает альбинизм у себя и своей сестры, а также упоминает, что звуки постоянно вызывали у него ощущение цвета, а буквы, цифры и дни недели имели «свои» цветовые обозначения [1].
Довольно часто синестезия встречается у людей творческих профессий: поэтов, композиторов и художников. Известный писатель Владимир Набоков был синестетом — по-видимому, как и его мать и сын. «Черно-бурую группу составляют: густое, без галльского глянца, А; довольно ровное (по сравнению с рваным R) Р; крепкое каучуковое Г; Ж, отличающееся от французского J, как горький шоколад от молочного; темно-коричневое, отполированное Я. В белесой группе буквы Л, Н, О, X, Э представляют, в этом порядке, довольно бледную диету из вермишели, смоленской каши, миндального молока, сухой булки и шведского хлеба» — так Набоков сравнивал два синестетических алфавита — русский и французский. Пример Набокова интересен тем, что синестезия передавалась по наследству в их семье, а значит, могла иметь генетические основы.
Само существование синестезии несколько озадачивает при первом знакомстве, но в дальнейшем оно становится всё более понятным. На самом деле мы все, так или иначе, имеем способность к переводу сигналов одной сенсорной системы на «язык» других чувств. Вот простой пример. На рисунке 1 изображены два существа; одно называют буба, а другое — кики. Попробуйте угадать, кого как зовут. Подавляющее большинство людей решает этот вопрос так: буба обладает округлыми очертаниями, а существо с острыми углами — это кики. Более мягкое и плавное звучание слова «буба» связано у нас с мягкими и плавными переходами этой фигуры, а угловатое существо кики хорошо совпадает с грубым и отрывочным произношением его названия.

Рисунок 1. Буба и кики. Два существа практически безошибочно определяются из-за врожденных способностей людей к переводу одних ощущений в другие.
Путаница в голове
Существуют две основные теории, объясняющие возникновение синестезии. Первая теория — теория кросс-активации — предполагает, что при возникновении сигнала в участке коры, «отвечающем» за конкретное чувство, возбуждение нервных клеток передается на соседние отделы [2]. Это может происходить за счет связей между нейронами, которые у людей без синестезии были уничтожены во время прунинга — процесса избавления нейронов от ненужных, избыточных связей с другими нейронами, который начинается в некоторых отделах нашего мозга незадолго до рождения, в дальнейшем охватывает всё больше его частей, а заканчивается к половому созреванию. Нейроны изначально очень щедро раскидывают свои связи, а затем происходит своеобразный естественный отбор среди нервных окончаний (рис. 2). Главная основа такого отбора — правило «пользуйся или откажись» («use it or lose it»). Чем реже используется какой-то отросток нейрона, тем меньше у него шансов «выжить» в ходе прунинга. Нейрон как бы втягивает эти малоиспользуемые отростки и дальше не тратит свои ресурсы на поддержание их работы. Возможен такой вариант развития событий, что в некоторых участках мозга прунинг не произошел так, как нужно, и избыточные связи продолжили свое существование. Эти дополнительные нейронные «тропинки» и обеспечивают возможность передачи возбуждения на соседние участки коры.
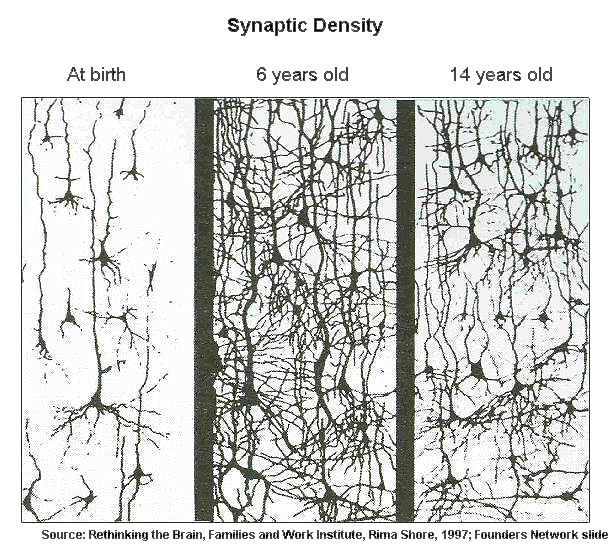
Рисунок 2. «Use it or lose it». С момента рождения в некоторых отделах мозга количество связей между нейронами постепенно увеличивается, но со временем тенденция меняется — нейроны начинают избавляться от лишних контактов.
Теорию кросс-активации можно продемонстрировать на основе анализа графемно-цветовой синестезии — феномена, при котором буквы и цифры вызывают ощущение цвета. Ключевым участком зрительной коры при определении цвета является область V4. Сама область V4 находится возле участка веретеновидной извилины, функция которого заключается в идентификации слов и букв (visual word form area, VWFA) (рис. 3). Если VWFA будет активироваться при восприятии букв, то у синестетов нервный сигнал по дополнительным каналам будет переходить в зону V4. Автоматическая активация этой области приведет к возникновению ощущения цвета. Эти теоретические предположения подтверждаются при исследованиях мозга синестетов с помощью фМРТ — функциональной магнитно-резонансной томографии (рис. 4) [3].
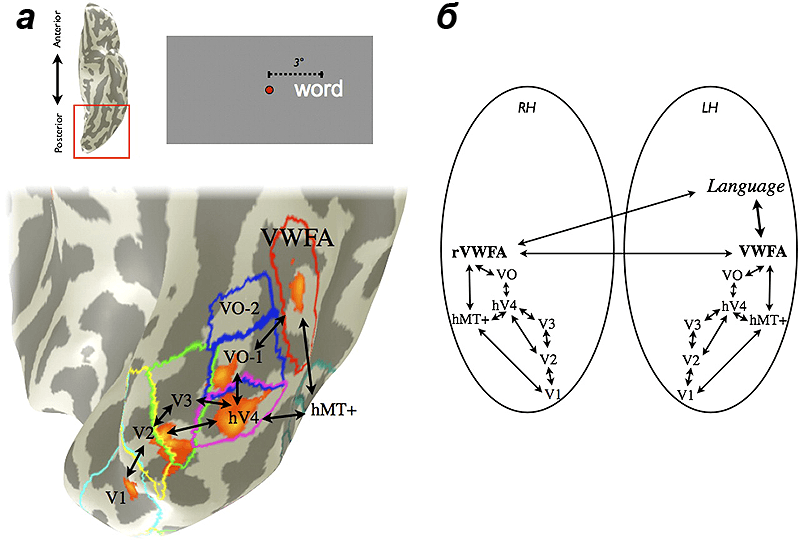
Рисунок 3. Расположение различных функциональных зон коры головного мозга. Вся визуальная информация изначально попадает в первичную зрительную кору (V1). Затем нюансы изображения распознаются в других участках (V2-4). Распознавание цвета происходит в области V4, которая может спонтанно активироваться при передаче сигнала из близлежащей зоны VWFA, которая отвечает за распознавание букв.
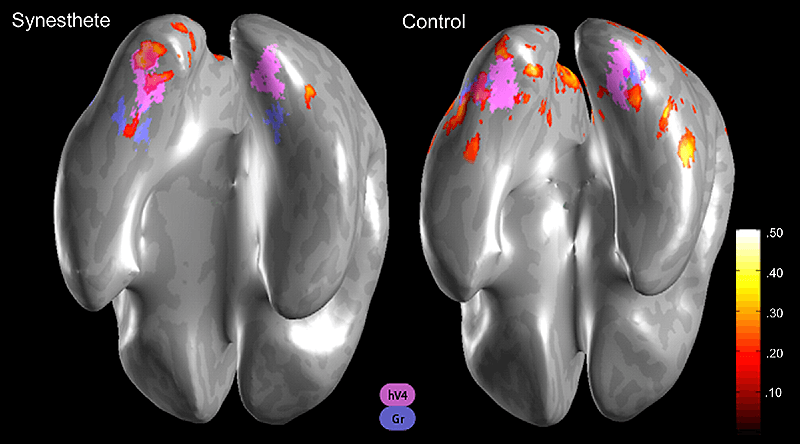
Рисунок 4. Активация области V4 у синестетов при восприятии графических символов. На снимках фМРТ заметно, что при выполнении этой задачи у синестетов область V4 (обозначена розовым) активна — в отличие от испытуемых без способностей к синестезии.
Вторая теория, объясняющая феномен синестезии, основывается на довольно интересном явлении. У человека нервные сигналы поступают от органов чувств в первичные корковые зоны, где происходит их восприятие. Оттуда информация направляется в ассоциативные области коры, где распознаются образы. Хорошим примером может служить зрительное восприятие. Мы смотрим фильм, и поток света попадает нам на сетчатку. Оттуда зрительная информация в виде электрических сигналов попадает в первичную зрительную область затылочной коры. В дополнительных сенсорных областях затылочной коры происходит распознавание цвета и формы увиденных нами объектов. В дополнительных областях височной коры распознаются конкретные образы, а теменная кора формирует карту расположения объектов и анализирует направления их движения.
Обычно процесс передачи информации из первичных отделов коры в ассоциативные происходит только в одном направлении, но ученые предполагают, что у синестетов нервные сигналы могут двигаться и в обратном направлении. Это не означает, что возбуждение идет обратно по тому же нейрону. Сигнал, пришедший из первичных отделов коры в ассоциативные, возбуждает активность в нервных клетках этого отдела. Далее сигнал по отросткам активированных нейронов снова устремляется в первичные отделы коры. Кроме этого, сигналы могут вернуться не в свое первичное поле, а «заглянуть» в другие участки коры, тем самым формируя дополнительные ощущения. Причиной может стать недостаточная работа ГАМКергической системы, подавляющей излишнюю активность в головном мозге [4]. Эта система вовремя не перекрывает путь побочно возникающим сигналам, и они устремляются к различным сенсорным участкам коры, формируя не связанные с первичным стимулом ощущения. Нервное возбуждение ведет себя как первоклассник: получив взбучку от одного товарища, он начинает бегать по классу и задирать других, пока не достигнет желаемого — не заставит кого-то ему ответить. Похожий процесс происходит в мозге при приеме некоторых психоактивных веществ, например, ЛСД или кетамина. У людей, принявших эти препараты, возникают различные синестетические переживания: звуки и температурные стимулы могут вызвать визуальные образы, а определенные запахи — тактильные ощущения. Кроме того, прием психоделиков с серотонинергическим эффектом вызывает усиление уже существующей синестезии. По-видимому, в механизме синестезии, индуцированной приемом психоделиков, задействован серотонин, так как многие препараты этой группы обладают серотонинергической активностью [5].
Синестетическое наследство
Синестезия — это интересный феномен, и совершенно неясно, откуда он берется. Учитывая, что синестетические ощущения не определяются индивидуальным опытом человека, вероятнее всего, предпосылки к их появлению врожденные. А если это так, то у синестезии может быть генетическая основа. В 90-х годах XX века считалось, что способность к синестезии может быть связана с X-хромосомой, а ее наследование носит доминантный характер. В дальнейшем стало ясно, что механизмы наследования несколько сложнее [6].
Классическим способом исследования генетических основ того или иного состояния является генеалогический метод — составление семейного дерева с указанием на нём тех родственников, которые имели исследуемый признак. В 2008 году в журнале Cognition была опубликована работа, исследующая наследуемость синестезии генеалогическим методом. У 42% из 53 участвовавших в ней синестетов были обнаружены родственники первой степени родства (родители или дети), у которых тоже отмечалась способность к синестезии. Также оказалось, что в одной семье могут сосуществовать разные типы синестезии с разной силой выраженности [7]. Синестезия передается членам семьи обоих полов, что указывает на возможность расположения «гена синестезии» на X-хромосоме или на любой аутосоме. Кроме того, в возникновении феномена могут играть роль факторы среды.
Другой метод изучения наследуемости признака — близнецовый. Близнецы удобны для генетических исследований тем, что достоверно имеют общие гены. В случае монозиготных (однояйцевых) близнецов число общих генов равно 100%, а в случае гетерозиготных — в среднем 50%. Цель близнецовых исследований — поиск конкордантности, то есть степени совпадения признака у обоих близнецов. Высокая конкордантность у монозиготных близнецов при более низкой у гетерозиготных указывает на значительное влияние генетического компонента при формировании исследуемого признака. Если в обеих группах близнецов конкордантность низкая, то это позволяет сделать вывод о преимущественном влиянии окружающей среды на формирование признака. Американские ученые сравнили конкордантность синестезии у 34 пар близнецов — 23 гомозиготных и 11 гетерозиготных, — в которых хотя бы один из близнецов имел графемно-цветовую синестезию [8]. Оказалось, что у гомозиготных близнецов конкордантность синестезии составляет 73,9%, а у гетерозиготных — 36,4% (рис. 5). При более детальном исследовании гетерозигот ученые обратили внимание на то, что в разнополых парах близнецов синестезия у обоих близнецов встречается реже, чем в однополых. По мнению исследователей, это может указывать на значительную роль среды при зарождении способности к синестезии.
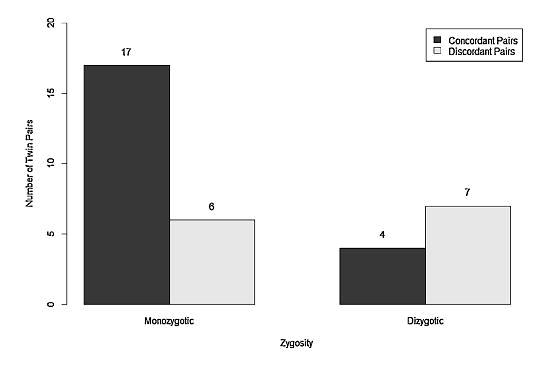
Рисунок 5. Конкордантность синестезии у гомо- и гетерозиготных близнецов. На графике видно, что синестезия встречается у обоих гомозиготных близнецов чаще, чем у гетерозиготных, что указывает на значительное влияние генов в развитии этого феномена. Однако неполная конкордантность у гомозиготных близнецов говорит о важности средовых факторов для развития этого состояния.
Следующий этап генетических исследований — непосредственная работа с последовательностью нуклеотидов, то есть изучение структуры ДНК. В работе [9] анализировали геномы 196 человек из 43 семей, включенных в базу данных Кембриджской группы по изучению синестезии (Cambridge Synaesthesia Research Group). Из этого числа 121 человек имел звуко-цветовую синестезию, то есть ощущение цвета у них вызывалось восприятием звукового стимула (явление очень похоже на графемно-цветовую синестезию). При полногеномном сканировании обнаружились участки ДНК, которые больше всех прочих были связаны с синестетическими переживаниями: участки на длинных плечах 2-ой и 5-ой хромосом, а также на коротких плечах 6-ой и 12-ой. Это исследование не выявило связи синестезии с Х-хромосомой. В обозначенных участках локализованы гены, влияющие на развитие и работу нервной системы человека. Например, участок 5-ой хромосомы содержит ген DPYSL3, вовлеченный в процессы нервной пластичности и аксонального роста. Этот ген особенно активен в последние недели перед и в первое время после рождения. На выявленном участке 6-ой хромосомы находятся гены, отвечающие за работу со звуковой информацией. Мутации в этих генах приводят к дислексии, однако не исключено, что не столь драматические изменения могли вызвать синестезию. Другое исследование выявило связь графемно-цветовой синестезии в нескольких семьях с определенной областью 16-ой хромосомы, в которой находятся: ген, связанный с передачей сигнала от нейрона к нейрону (GABARAPL2), ген, вовлеченный в развитие мозга (NDRG4) и ген, имеющий отношение к упомянутому выше прунингу (KATNB1) [10].
Как можно заметить, перечисленные работы описывают генетику того типа синестезии, который можно объяснить кросс-активацией — «нечаянным» распространением сигнала из одного отдела коры в другой, расположенный неподалеку. Несколько смущает разнородность данных по генам, которые вовлечены в этот тип синестезии, но этому можно найти объяснение. Развитие мозга — это сложное явление, которое состоит из множества процессов. За каждый из них отвечает несколько генов, которые, вероятнее всего, выстраивают целые функциональные цепи, а может быть, даже и сети. Нефатальные нарушения в работе генов этих сетей могут менять всю работу нейрона, приводя к возникновению отдельного эндофенотипа работы коры. Главная особенность синестетического эндофенотипа — гиперсвязанность одних отделов мозга с другими, и эта гиперсвязанность может определяться изменением работы генетической сети в целом, а не нарушением активности конкретного гена. Синестезия — это не болезнь, а то состояние мозга, которое еще раз показывает, насколько сложна и восхитительна жизнь этого удивительного органа.
Источник