ЭВОЛЮЦИЯ ОРГАНОВ ЧУВСТВ
Большинство животных чувствуют окружающую среду– одноклеточныечувствуют органоидами, многоклеточные –нервными клетками или специальными органами.
У простейших осязание поверхностью мембраны у амебы, ресничками у инфузории, у эвглены светочувствительный глазок, химическое восприятие солености воды у инфузории.
У гидрыраздражение воспринимают нервные клетки и передают возбуждение кожно-мускульным клеткам, они и вызывают сокращение тела гидры.
Плоские свободноживущие черви имеют простые глаза, органы осязания и равновесия. У круглых червей слабо развиты органы чувств: на головном конце тела — чувствительные сосочки и мелкие щелевидные углубления – органы химического чувства.
Кольчатые черви имеют чувствительные клетки, разбросанные по всему телу, а специализированных органов чувств нет.
У двустворчатых моллюсков органы чувств развиты слабо, есть органы равновесия у основания ноги, химического чувства в виде клеток по всему телу.
У брюхоногих моллюсков развиты глаза и органы осязания, у головоногих моллюсков глаза сложно устроены и аналогичны глазам млекопитающих;
Глаза раков– сложные фасеточные из 3 тыс. простых глазков, а зрение мозаичное, т.е. каждый глазок видит только часть предмета, органы равновесия в усиках, а органы осязания и химического чувства в виде волосков и щетинок на разных частях тела.
У пауков органы зрения представлены простыми и сложными глазами, органы осязания — волосками на конечностях, органы химического чувства в углублениях покровов тела.
Насекомыеобладают простыми и фасеточными глазами, способны видеть ультрафиолетовую часть спектра, распознают цвета, но видят хорошо только на близком расстоянии, сложно устроены органы равновесия, обоняния, осязания, вкуса и слуха (тимпанальные органы в виде тонкой перепонки) .
Ланцетник, как представитель хордовых, имеет примитивные светочувствительные глазки вдоль нервной трубки, обоняние – углубление на переднем конце нервной трубки, осязательные клетки на щупальцах рта.
У рыбнаоборот развиты органы зрения, роговица плоская, а хрусталик круглый эти особенности дают более дальнозоркое зрение, развит слух, обоняние, органы вкуса. Боковая линия воспринимает направление и силу тока воды.
У земноводных из-за выхода на сушу органы чувств усложнились: глаза с выпуклой роговицей и линзовидным хрусталиком, есть 3 века (верхнее, нижнее, мигательная перепонка в углу глаза) – защита от высыхания и загрязнения; появилось среднее ухо, органы обоняния – наружные и внутренние ноздри.
У пресмыкающихся глаза снабжены тремя веками, а змеи и гекконы имеют сращенные верхнее и нижнее веки, они становятся прозрачны, органы слуха более приспособлены к восприятию звуковых волн, есть внутреннее и среднее ухо; осязание в виде чувствительных клеток с волосками на чешуйках тела, обоняние и вкус расположены в области ротовой полости, есть орган термического чувства в виде ямок с чувствительными клетками между глазами и ноздрями.
У птиц слабо развито обоняние (кроме падальщиков), слух представлен внутренним и средним ухом, а улавливать звук помогают кроющие перья вокруг барабанной перепонки. Глазаптиц прекрасно приспособлены к рассматриванию объектов, острота зрения очень большая (сокол различает добычу на расстоянии более 1000м.). Глаза очень велики, у сов они по объему равны глазу взрослого человека и в 5 раз больше массы мозга, у хищных птиц, высматривающих добычу на большом расстоянии, глаза в 3 раза превышают массу мозга — это позволяет получать большие размеры изображения на сетчатке и тем самым яснее различить его детали, защищены тремя веками.
Аккомодация достигается двойным путем:
1)изменением формы хрусталика под действием ресничной мышцы,
2) изменением расстояния между хрусталиком и сетчаткой, под действием кольцевых мышц, окружающих склеру и меняющих форму глазного яблока. Большинство птиц имеют монокулярное зрение (каждый глаз видит предметы отдельно), бинокулярным (позволяющим видеть предметы двумя глазами одновременно) обладают совы, все обладают цветным зрением, на сетчатке есть участок максимально острого зрения – желтое пятно (ямка), у питающихся подвижными объектами дневных хищников, цапель, ласточек два желтых пятна, у стрижей только одно, поэтому их способы ловли добычи на лету менее разнообразны. Очень подвижный зрачок предотвращает излишнюю засветку сетчатки при быстрых поворотах.
У млекопитающих органы чувств по — разному развиты.. На первом месте зрение для обитателей открытых пространств, обоняние и слух – для ночных и животных лесов, обитателей водоемов. Они обладают бинокулярным зрением. Цветное зрение сравнительно с птицами развито слабо. Все цвета спектра различают только люди и высшие обезьяны, полевка видит красный и желтый цвет, в тоже время они распознают особенности формы, детали предметов, движения, позу и мимику. Это обеспечено усложнением зрительного анализатора в коре головного мозга. Для зверей дневного образа жизни и обитателей открытых пространств, большая часть информации идет через зрительный канал, глаза копытных и грызунов по бокам головы, что дает круговой обзор- это важно для постоянно ожидающих нападения врагов. Обитатели лесов и зарослей имеют менее развитое зрение, у норников (кротов, слепышей) глаза перестают функционировать, зарастая кожей. У обезьян, кошачьих есть необходимость точного определения расстояния, поэтому их глаза расположены фронтально, что увеличивает поле бинокулярного зрения. У кошачьих и копытных между склерой и сосудистой оболочкой есть слой клеток с кристалликами (зеркальце), обуславливающее «свечение» глаз отраженным светом, что улучшает видение ночью.
Наземные млекопитающие в органе слуха имеют внутреннее, среднее и наружное ухо (наружный слуховой проход и ушную раковину). Органы обоняния чуткие, они расположены в обонятельных отделах носовой полости, орган вкуса лучше развит у травоядных животных, осязание представлено рецепторами в коже, имеются особые жесткие волосы – вибриссы, их корни связаны с окончаниями нервов, особенно развиты вибриссы на морде.
Источник
Эволюция органа зрения
Орган зрения, как и все другие органы чувств, в ходе филогенетического развития претерпел сложную эволюцию, которая шла в сторону большего и лучшего приспособления глаза к восприятию окружающего мира.
Простейшей формой зрения следует считать начало реакции на свет. Почти все живое чувствительно к свету. У растений световая реакция проявляется гелиотропизмом (например, головка цветущего подсолнечника в течение всего дня повернута к солнцу).
Простейший орган зрения у дождевого червя представляет собой отдельные светочувствительные клетки, расположенные изолированно в эпидермисе и способные различать только свет и его направление.
Так называемые зрительные клетки пиявки сконцентрированы в определенных местах, объединены в группы по 5-6 штук, с внутренней стороны ограничены прослойкой темного пигмента. В глазах морской звезды обнаружена начальная структура нейроэпителия. Его световоспринимающие концы обращены к свету, нервные волокна собраны в широкий тяж, которые можно считать примитивными нервами. Наружная часть глаза имеет форму ямки, выстланной сверху покровным эпителием. Глаз кольчатых червей еще более сложен. Он имеет вид круглой полости, содержащей прозрачную массу — своеобразное стекловидное тело. Между чувствительными клетками находятся клетки пигментного эпителия, появляются вставочные клетки, что соответствует опорной глиозной ткани сетчатки высших животных. Если глаза простейших живых существ реагируют только на свет и изменение его интенсивности, то более развитые глаза способны формировать образ.
У моллюска, стоящего еще на достаточно низкой ступени развития, глаз напоминает таковой высших животных. Клетки нейроэпителия направлены не к свету, не к центру глаза, а от света.
Таким образом, возникает тип перевернутой сетчатки, что характерно для глаз высших животных. В глазу моллюска уже есть подобие линзы. Фоторецепторы скрываются в углублениях, где защищены от яркого света, уменьшающего способность улавливать движущуюся тень. Здесь линза выполняет функцию прозрачной защитной мембраны. Далее постепенно начинает совершенствоваться защитный аппарат глаза.
Сложно организованные глаза часто бывают при простом мозге. У некоторых членистоногих, включая насекомых, имеются сложные фасеточные глаза, содержащие более тысячи фасеток. Подобные глаза являются специальными факторами движения.
Глаз человека по своей структуре — типичный глаз позвоночных. Однако он имеет существенные функциональные отличия.
Глаз высших животных развивается из разных тканевых источников. Сетчатка и зрительный нерв формируются из эктоневральной закладки центральной нервной системы. На 2-й неделе эмбриональной жизни, когда мозговая трубка еще не замкнута, на дорсальной поверхности медуллярной пластинки появляются два углубления — глазные ямки. На вентральной поверхности им соответствует выпячивание. При замыкании мозговой трубки ямки перемещаются, принимают боковое направление. Эта стадия носит название первичного глазного пузыря. На этой стадии развития глаза полость мозга свободно переходит в полость первичного глазного пузыря. Вершины глазных пузырей почти вплотную подходят к эктодерме; их разделяет лишь узкий слой мезодермы. Такие соотношения выявляются на 3-й неделе, когда длина всего зародыша составляет 3 мм. С конца 4-й недели развития возникает хрусталик. Вначале он имеет вид утолщения покровной эктодермы в том месте, где первичный глазной пузырь начинает превращаться во вторичный. Первичный пузырь растет неравномерно; отмечается быстрый рост задних и боковых стенок, в то время как рост передних и нижних стенок задерживается. Быстрорастущие задние и боковые области образуют передние и нижние части. Однослойный первичный глазной пузырь на полой ножке превращается во вторичный пузырь, состоящий из двух слоев, — глазной бокал. При образовании глазного бокала возникает зародышевая щель, которая заполняется прилежащей мезодермой. Между зачатком хрусталика и внутренней стенкой бокала остается небольшое количество мезенхимальных клеток, из которых формируется первичное стекловидное тело).
В этот период развития хрусталик занимает почти всю полость глазного яблока. Очень интенсивно происходит размножение клеток, выстилающих верхнюю стенку хрусталикового пузырька. Постепенно вытягиваясь, они заполняют всю полость пузырька. Край глазного бокала снизу начинает все больше ввертываться, формируя вторичную зародышевую щель. Через эту щель проникает большое количество мезенхимы, которая образует богатую сосудистую сеть стекловидного тела. Вокруг хрусталика формируется сосудистая капсула. В возрасте 6 недель зародышевая щель глаза и зрительного нерва закрывается, начинает дифференцироваться ножка глазного бокала, образуется a. hyaloidea, питающая стекловидное тело и хрусталик.
Наружный листок бокала в дальнейшем превращается в пигментный слой сетчатки, из внутреннего же развивается собственно сетчатка. Края глазного бокала, прорастая впереди хрусталика, образуют радужную и ресничную части сетчатки. Ножка, или стебелек, глазного бокала удлиняется, пронизывается нервными волокнами, теряет просвет и превращается в зрительный нерв. Из мезодермы, окружающей глазной бокал, очень рано начинают дифференцироваться сосудистая оболочка и склера. В мезенхиме, которая прорастает между эктодермой и хрусталиком, появляется щель — передняя камера. Мезенхима, лежащая перед щелью, вместе с эпителием кожи превращается в роговицу, лежащая сзади, — в радужку. К этому времени начинается постепенное запустевание сосудов стекловидного тела. Сосудистая капсула хрусталика атрофируется. Внутри хрусталика образуется плотное ядро (зародышевое ядро хрусталика), объем хрусталика уменьшается. Стекловидное тело приобретает прозрачность.
Веки развиваются из кожных складок. Они закладываются кверху и книзу от глазного бокала, растут по направлению друг к другу и спаиваются своим эпителиальным покровом. Спайка эта исчезает к 7 месяцам развития.
Слезная железа возникает на 3-м месяце развития, слезный канал открывается в носовую полость на 5-м месяце. К моменту рождения ребенка весь сложный цикл развития глаза не всегда оказывается полностью завершенным. Обратное развитие элементов зрачковой перепонки, сосудов стекловидного тела и хрусталика может происходить в первые недели после рождения.
Т. Бирич, Л. Марченко, А. Чекина
«Эволюция органа зрения» статья из раздела Офтальмология
Источник
Откуда пошло зрение
В геноме гидры — кишечнополостного животного — открыт новый класс опсинов — белков, реагирующих на свет
Автор
Редакторы
Противники теории эволюции часто заявляют, что случайные изменения способны лишь разрушить какую-либо функцию организма, но не создать новую. Однако начинают появляться исследования, аккуратно показывающие, как именно могли возникнуть новые системы в процессе развития живого мира. Мало того: учёным, похоже, удалось «отследить» возникновение зрения, часто приводимого креационистами как пример системы, возникновение которой в результате последовательного развития невозможно.
Противники идеи эволюции часто превратно толкуют Дарвина, который якобы и сам признавал, что «трудно поверить в то, что столь сложный живой прибор создан в ходе естественного отбора путем проб и ошибок». Но на самом деле «отец» эволюционной концепции писал другое: «Утверждение, что столь совершенный орган, как глаз, мог возникнуть путем естественного отбора, способно поразить всякого. Тем не менее, если мы знаем для любого органа длинный ряд градуальных усложнений, из которых каждое полезно для его обладателя, то при меняющихся условиях жизни приобретение путем естественного отбора любой возможной степени совершенства органа логически вполне возможно. В тех случаях, когда нам неизвестны промежуточные или переходные стадии, мы должны быть весьма осторожны в заключении, что они никогда и не существовали, так как метаморфозы многих органов показывают, какие удивительные изменения функции оказываются, по крайней мере, возможными».
Учёные из университетов Санта-Барбары в Калифорнии и Квинсланда в Австралии провели подробное филогенетическое изучение опсинов — белков, отвечающих за рецепцию света в животных. (Зрение животных основано на светочувствительном кофакторе ретинале, в то время как фототаксис растений и грибов базируется на других молекулах.) В основе исследования лежит идентификация нового класса опсинов (названных книдопсами) в древнем примитивном классе гидроидных — многоклеточных животных, принадлежащих к типу кишечнополостных (Cnidaria). Анализ геномов эволюционно предшествующих гидрам животных — губок (тип Spongia) — выявил, что в них не встречается белков, родственных каким-либо известным формам фоторецепторов.
«Мы не только первыми исследовали гены, отвечающие за зрение в этих древних животных. То, что мы не обнаружили их в предшествующих жизненных формах — губках, — позволяет нам поставить точку отсчёта эволюционного развития зрения у животных», — говорит Дэвид Плачецкий (David Plachetzki), первый автор статьи, опубликованной в журнале PLoS ONE [1].
Свежеоткрытая группа опсинов дополняет уже известные ветви этих светочувствительных белков, отвечающих за зрение в различных формах животного мира:
- Зрение позвоночных основано на рецепторных клетках в виде «ресничек», образующих сетчатку глаза. В мембранах этих клеток (палочки и колбочки у животных с цветным зрением) расположен родопсин[2], связывающийся с Gi/t α-субъединицей G-белкá трансдуцина;
- Сложный глаз членистоногих (в частности, насекомых) содержит стержневидные клетки со складчатой мембраной, светочувствительный пигмент которых связывает Gq α-субъединицу;
- Go-опосредованный вид фоторецепции (у некоторых моллюсков и др.) действует через активацию Go α-субъединицы G-белкá.
Открытие новой группы опсинов позволило увеличить «разрешение» филогенетического анализа, который проводился и ранее. Проанализировав родство белков этих четырёх групп фоторецепторов, учёные пришли к выводу, что максимальное различие наблюдается между группой «реснитчатых» опсинов и группой, в которую попадают все остальные белки. Приняв в качестве рабочей гипотезы монофилию всех опсинов (то есть то, что все эти белки имели общего эволюционного предка), они объясняют возникновение этих групп с помощью двух последовательных дупликаций генов белка-прародителя. (Дупликация генов на настоящий момент считается одним из наиболее вероятных механизмов белковой эволюции. Согласно нему, ген «раздваивается» в геноме, и каждая его копия продолжает эволюционировать независимо, иногда приобретая новые функции.) По приблизительным оценкам, общий «предок» всех опсинов существовал 600 млн. лет назад (рис. 1).
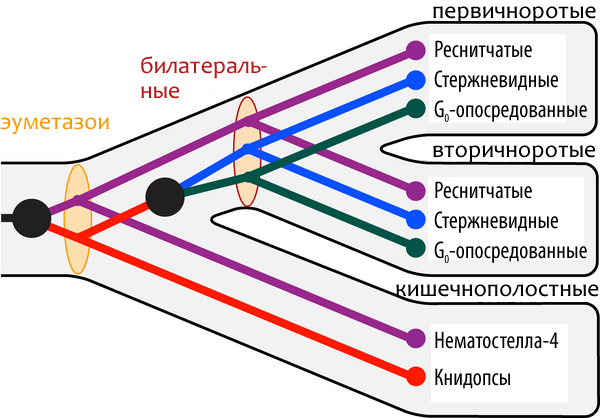
Рисунок 1. Гипотеза эволюции опсинов в рамках развития животного царства. Чёрными точками показаны моменты, в которые происходила дупликация генов (и, следовательно, число независимых белковых групп в одном организме увеличивалось). Овалами показаны «развилки» в эволюции фоторецепторов, сопровождающие появление новых групп животных. Согласно этой гипотезе, и «реснитчатые» опсины, и книдопсы существовали в эуметазойном предшественнике кишечнополостных и билатерально-симметричных животных. А вот «стержневидные» и G0-опосредованные формы появились уже в процессе эволюции билатеральных животных, но не раньше.
Кстати, реснитчатые опсины обнаружены и у кишечнополостных — но не у гидры, а у нематостеллы. Первая, видимо, утратила их в процессе эволюции.
Кроме того, анализ эволюционной вариабельности четвертой цитоплазматической петли опсинов показал, что участок, ответственный за связывание G-белкá, эволюционирует одновременно с «переключением» фоторецептора на нового G-белкóвого «партнёра».
Профессор Тодд Окли (Todd Oakley), заведующий лабораторией в Университете Санта-Барбары (и ведущий автор статьи), говорит, что до их работы практически не было исследований, в которых аккуратно демонстрировалось бы, что эволюционные изменения белка сопровождаются появлением новой функции. В данном же случае на лицо образование новых взаимодействий с G-белкáми, обнаруженными не только в многоклеточных, но и одноклеточных животных, а также в растениях и грибах. Такое новое белок-белковое взаимодействие можно считать фундаментом нового функционального механизма зрения, который, как уже было сказано, в разных группах животных основывается на различных биохимических процессах.
«Наветы противников эволюции просто-напросто неверны, — говорит Окли. — В нашей статье показано, что специфические мутации в продублированных генах опсинов задают новые, ранее не существовавшие межбелковые взаимодействия. Эти взаимодействия определяют работу всего механизма зрения, который отличается в разных группах животных».
Что пока остаётся невыясненным — это зачем же, всё-таки, гидрам опсины. Гидры — хищники, и можно было бы сказать, что светочувствительные пигменты требуются им для выслеживания жертвы, тем более что эти белки экспрессируются в области рта животного (показано тёмно-синим на заглавной картинке). Однако специальных органов зрения у гидр нет, хотя все биохимические пути, необходимые для рецепции света — присутствуют.
Источник