Анатомия и физиологи .
1.3. Действие электрического тока на возбудимые ткани. Пассивные и активные сдвиги мембранного потенциала. Законы раздражения возбудимых тканей.
В натуральных условиях причиной возникновения потенциала действия (ПД) является местный ток, который возникает между возбужденным и покоящимся участками мембраны. Поэтому электрический ток является адекватным раздражителем для возбудимых тканей.
В процессе возбуждения ткани в ней происходят сдвиги мембранного потенциала, вначале они носят пассивный характер, а затем активный.
Пассивные сдвиги потенциала возникают в ответ на действие любого раздражителя за счет его энергии. В отчетливой форме они лучше всего выявляются при действии полюсов постоянного тока (катода и анода). Пассивные изменения мембранного потенциала развиваются в предпиковом периоде потенциала действия (ПД), т.е. от момента нанесения раздражения до возникновения местного возбуждения.
Рассмотрим пассивные сдвиги мембранного потенциала, возникающие под катодом и анодом. Это явление называется физическим электротоном : соответственно физическим кат- и анэлектротоном .
Как известно, живая ткань является проводником электрического тока второго рода, и в ней электрические силовые линии проходят от анода к катоду. Там, где силовые линии входят в ткань через наружные электроды, формируется положительный заряд, где выходят из ткани – отрицательный . Таким образом, в месте приложения анода на наружной поверхности мембраны увеличивается положительный заряд, происходит как бы дозарядка емкости мембраны. Такое явление называется пассивной гиперполяризацией. В месте приложения катода выходящие силовые линии формируют на наружной поверхности мембраны отрицательный заряд, происходит как бы разрядка емкости мембраны. Такое явление называется пассивной деполяризацией. Объясняются эти явления тем, что мембрана – это «тканевой» конденсатор, наружная и внутренняя её поверхности являются обкладками, а слой липидов – диэлектриком. Но в мембране, в отличие от идеального конденсатора, имеются каналы (неселективные каналы), через которые происходит утечка ионов, и поэтому ее сопротивление не равно бесконечности, а имеет определённое значение. Поэтому мембрану сравнивают с конденсатором, к которому параллельно включено сопротивление, и когда электрический ток проходит через эту цепь (ёмкость и сопротивление), то через сопротивление происходит утечка зарядов. Произведение ёмкости мембраны (С) на её сопротивление (R) называется постоянной времени мембраны , она определяет скорость сдвига мембранного потенциала; чем она меньше, тем быстрее нарастает потенциал. Поскольку изменения мембранного потенциала при действии подпорогового постоянного тока (до 50% от порогового уровня) не связаны с активацией селективных каналов мембраны и изменением их ионной проницаемости, постольку их называют пассивными, или электротоническими.
Электротонические потенциалы:
1) нарастают и исчезают постепенно;
2) способны к суммации;
3) распространяются с декрементом;
4) при увеличении подпорогового тока до 50% от порогового значения возникает активный сдвиг потенциала в виде местного возбуждения.
Действие постоянного тока на возбудимые ткани изучал Пфлюгер (Германия, 1859 г). Он изложил закономерности его действия в виде полярного закона , который имеет три положения.
1. Постоянный ток действует на живую ткань раздражающим образом только в момент замыкания или размыкания цепи.
2. В момент замыкания возбуждение возникает под катодом, а в момент размыкания – под анодом. Под катодом возбудимость и проводимость ткани увеличиваются, а под анодом – уменьшаются; эти явления были названы соответственно физиологическими кат- и анэлектротоном и являются следствием физического электротона. Электротонические изменения постепенно ослабевают по мере удаления от электродов. Между электродами имеется индифферентная точка, в которой электротонические изменения отсутствуют.
3. Катодраздражающее действие постоянного тока сильнее, чем анодраздражающее.
Механизм этих положений стал понятен позже, когда были изучены свойства мембраны и ее каналов.
При замыкании цепи постоянного тока изменяется его градиент (скорость нарастания во времени), т.е. сила тока увеличивается от нуля до величины электродвижущей силы источника постоянного, а при размыкании цепи падает от величины ЭДС источника тока до нуля, и эти изменения электрического стимула во времени и обладают раздражающим действием. По существу, это закон градиента раздражения , который мы рассмотрим позже.
При замыкании цепи постоянного тока под катодом происходит деполяризация мембраны, которая носит пассивный характер электротонического возбуждения, если сила катодраздражающего действия равна 5-50% от пороговой величины. Установлено, что при этом происходит сдвиг исходного уровня МПП (Е0) в сторону Ек, при этом порог деполяризации ( Δ V = Е0 — Ек) уменьшается. Эти изменения вызваны усилением входящего потока Na + по каналам утечки, который превышает также увеличенный выходящий ток утечки для К + и Сl — . Скорость этой деполяризации будет зависеть от постоянной времени мембраны.
Под анодом происходит пассивная гиперполяризация, т.е. увеличивается исходный уровень потенциала (Е0) и Δ V увеличивается, что приводит к снижению возбудимости и проводимости в этой области. На мембране в это время увеличивается поток К+ наружу по каналам утечки и снижается движение Na + внутрь клетки.
При длительном действии полюсов постоянного происходят противоположные явления, которые, однако, не связаны с изменением Е0, а связаны с изменением Ек. Рассмотрим эти явления.
Явление снижения возбудимости нервного волокна при длительной и сильной деполяризации было открыто русским ученым И.Б.Вериго в 1889 г.
При длительном подпороговом действии катода Ек возрастает настолько, что порог деполяризации становится намного больше исходной величины, одновременно резко падает амплитуда ПД. Это явление с особой лёгкостью возникает в препаратах, альтерированных различными агентами, понижающими амплитуду ПД (KСl, новокаин, механическое повреждение и т.д.). На мембране в это время происходит инактивация натриевых каналов и повышение проводимости мембраны для ионов К + (активация калиевых каналов).
При длительном действии анода снижается Δυ и Ек приближается к значению Е0. Теперь оказывается достаточным одного выключения анодного тока, чтобы ПП, быстро восстанавливающийся к исходной величине, достиг Ек, вызвал появление ПД и тем самым создал условия для возникновения анодразмыкательного возбуждения. На мембране в это время снижается калиевая проводимость и ослабляется исходная натриевая инактивация.
Явления кат- и анэлектротона справедливо рассматривались классиками электрофизиологии как прообраз, модель процессов возбуждения и торможения, разыгрывающихся в условиях естественной деятельности нервной системы. В реальных условиях на принципиально том же механизме, что и катэлектротоническое повышение возбудимости, основаны такие явления, как следовое повышение возбудимости, облегчение проведения возбуждения в синапсе, повышение возбудимости нейрона под влиянием его бомбардировки подпороговыми афферентными импульсами и другие явления, обусловленные подпороговой деполяризацией мембраны. С другой стороны, катодическая депрессия играет роль в механизмах некоторых видов периферического и центрального торможений, основанных на чрезмерной по силе или длительности деполяризации мембраны: это явление пресинаптического торможения в ЦНС, торможение по механизму пессимума Введенского и т.д. Повышение Ек лежит в основе явлений аккомодации ткани к раздражителю.
Анэлектротоническое понижение возбудимости лежит в основе следового снижения возбудимости после ПД.
Если сила катодраздражающего действия равна 50-95% от порогового значения, то развивается активная реакция в виде местного возбуждения.
Как было указано выше, при увеличении силы раздражителя (более 50% от пороговой величины) пассивная деполяризация мембраны порождает активную реакцию, – к электротоническому потенциалу присоединяется дополнительная активная деполяризация мембраны, называемая местным возбуждением, или локальным ответом. Локальным он называется потому, что и в экспериментах, и в естественных условиях он далеко не распространяется (на 2-3 мм от места возникновения). При местном возбуждении изменения на мембране идут уже за счёт энергии, накопленной мембраной в потенциале покоя, а не за счёт энергии раздражителя.



При пороговом раздражении пассивная деполяризация (электротонический потенциал) и локальный ответ суммируются друг с другом, и когда эта сумма достигает критической величины, возникает потенциал действия. В гигантских аксонах кальмара на долю локального ответа при пороговой силе раздражения приходится от 20 до 30% общего сдвига мембранного потенциала.

2. Во время фазы быстрой деполяризации ткань становится совершенно невозбудимой, эта фаза называется абсолютной (эффективной) рефрактерностью ( refraktar , нем. – невосприимчивый). В этой фазе даже супермаксимальные раздражители не способны вызвать генерацию нового ПД. Эта фаза обусловлена невозможностью дополнительной активации натриевых каналов и увеличением проницаемости калиевых каналов.
3. После абсолютной невозбудимости происходит постепенное восстановление возбудимости до исходного уровня – это фаза относительной рефрактерности. Она соответствует быстрой реполяризации мембраны, в это время происходит восстановление исходного состояния натриевых каналов, поток Na + в клетку уменьшается, а поток К + из клетки увеличивается. В это время можно вызвать генерацию нового ПД, но для этого должен быть надпороговый раздражитель.
4. Во время отрицательного следового потенциала возбудимость повышается – это называется фазой экзальтации ( exaltatio , лат. – возбуждённое, восторженное состояние), или супернормальной возбудимости. В это время подпороговые раздражители могут вызвать возбуждение, т.к. пороговый потенциал снижен, поскольку потенциал покоя ближе к критическому уровню деполяризации, чем в состоянии покоя.
5. В тканях, где имеется следовая гиперполяризация, ей соответствует фаза субнормальной возбудимости (сниженной возбудимости), за счёт удаления потенциала покоя от критического уровня деполяризации.

Или, например, если нервное волокно обработать местными анестетиками (новокаин, кокаин), ядами, наркотиками, которые ослабляют входящий поток ионов Na+ в клетку, то амплитуда ПД начнёт зависеть от силы раздражителя, т.е. перестанет подчиняться закону «всё или ничего».
Закон времени .
Этот закон был изучен при исследовании мышц, нервов Гоорвегом (Голландия, 1892), Вейсом (Германия, 1901) и Лапиком (Франция, 1909).
Закон устанавливает зависимость между силой раздражителя и временем его действия: «Пороговая сила любого раздражителя обратно пропорциональна длительности его действия в определенных пределах». То есть, чем меньше по времени действует раздражитель, тем больше требуется его сила для формирования возбуждения (рис. 8).


где I – сила тока;
t – время действия тока;
a и b – постоянные величины.
Минимальная сила тока, которая способна вызвать возбуждение (порог раздражения) была названа Лапиком реобазой . Наименьшее время, которое должна действовать реобаза, чтобы вызвать возбуждение, называется главным полезным временем . Если ток действует дольше этого полезного времени, это не имеет значения (бесполезно) для возникновения ПД. При укорочении времени действия тока (меньше полезного времени) для получения ПД необходимо увеличивать силу тока. При очень коротких стимулах восходящая часть гиперболы становится параллельной оси ординат, это означает, что при очень коротких раздражающих стимулах ПД не возникает, как бы ни была велика сила тока. Это явление используется в физиотерапии, при лечении больных токами УВЧ (ультравысокой частоты), для глубокого прогревания тканей.
Точное определение главного полезного времени практически трудно, т.к. величина реобазы непрерывно претерпевает колебания, связанные с колебаниями функционального состояния мембраны в покое, и точка перехода может смещаться влево или вправо, поэтому Лапик предложил измерять условную величину в качестве константы времени раздражения и назвал её «хронаксия». Хронаксия – это дополнительное полезное время – время, которое должен действовать ток, равный по величине удвоенной реобазе, чтобы вызвать возбуждение.
В неврологической практике проводят хронаксиметрию, чтобы установить поражение двигательных нервов. Если нерв повреждён или погибли мотонейроны спинного мозга (полиомиелит), то нервные волокна перерождаются и раздражающий стимул выявляет увеличение реобазы и хронаксии.
Закон градиента.
Для того чтобы раздражитель вызвал генерацию ПД, его сила должна нарастать достаточно быстро. При какой-то минимальной крутизне нарастания интенсивности раздражения (минимальный градиент) ответы на это раздражение исчезают, т.к. в ткани развивается процесс аккомодации ( accomodation , англ. – приспособление). Величина минимального градиента, выраженного в единицах реобаза в секунду (МА), является показателем скорости аккомодации. Чем круче должен нарастать ток, чтобы вызвать возбуждение, тем выше скорость аккомодации.
В основе механизма аккомодации лежат инактивация Na + -каналов и повышение калиевой проводимости, развивающиеся при «вкрадывающемся» в ткань токе.
Параметры возбудимости ткани.
Для оценки степени возбудимости тканей используют следующие критерии:
1) порог раздражения;
2) хронаксия;
3) уровень лабильности (максимальное количество импульсов, которое может воспроизвести ткань без трансформации ритма);
4) скорость аккомодации.
Источник
9. Законы раздражения. Полярный закон. Физиологический электротон. Като-дическая депрессия.
Полярный закон раздражения (закон Пфлюгера)

При замыкании цепи постоянного электрического тока возбуждение возникает только под катодом, а при размыкании — только под анодом
Доказать правильность закона можно при помощи опыта, в котором меняется расположение катода и анода на поврежденном и неповрежденном участках нерва, иннервирующего мышцу
Прохождение постоянного электрического тока через мембрану вызывает изменение мембранного потенциала покоя.
Так, при замыкании цепи возле катода скапливается его «-» заряд, который уменьшает «+» заряд наружной поверхности мембраны.
Разность потенциалов (между наружной и внутренней поверхностями мембраны) уменьшается, и мембранный потенциал изменяется в сторону уровня критической деполяризации, т.е. формируется деполяризация(возбудимость повышается).
Достижение критического уровня приводит к возникновению пикового потенциала (потенциала действия)
При замыкании цепи возле анода скапливается его «+» заряд.
Он увеличивает «+» заряд мембраны и величину мембранного потенциала
Мембранный потенциал удаляется от критического уровня, превышает значение потенциала покоя и формируетгиперполяризацию (возбудимость уменьшается)
При размыкании цепи, прекращение поступления дополнительного «+» заряда от анода приводит к уменьшению(восстановлению) заряда наружной поверхности мембраны.
Мембранный потенциал, уменьшаясь, приближается к критическому уровню
Формируется деполяризация (возбудимость увеличивается).
После достижения критического значения развивается пиковый потенциал
(Поскольку размыкание происходит после замыкания, а, следовательно, на фоне гиперполяризации и пониженной возбудимости, то для возникновения ПД необходим раздражитель, превышающий по силе пороговый — это анодно-размыкательный эффект)
При размыкании возле катода прекращается накопление его «-» заряда
Заряд наружной поверхности мембраны увеличивается (восстанавливается), мембранный потенциал, увеличиваясь, удаляется от критического уровня и возбуждение не возникает
Закон физиологического электротона
Действие постоянного электрического тока на ткань сопровождается изменением ее возбудимости
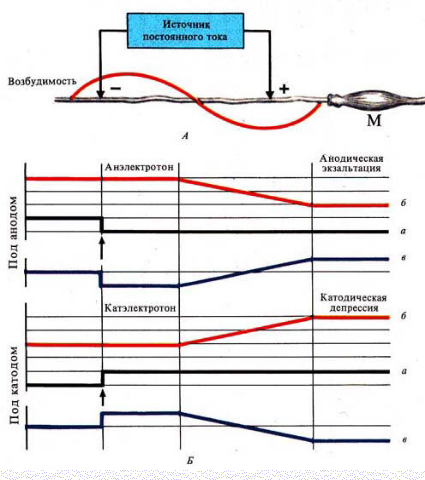

Различают 3 вида физиологического электротона или изменения возбудимости:
Катэлектротон — изменение возбудимости под катодом.
В момент замыкания под катодом формируется деполяризация и возбудимость повышается.
По мере удаления от катода количество его «-» зарядов, а следовательно и выраженность деполяризацииуменьшается.
В результате возбудимость уменьшается, но она остается выше, чем в состоянии покоя
Анэлектротон — изменение возбудимости под анодом.
В момент замыкания под анодом формируется гиперполяризация и возбудимость снижается.
По мере удаления от анода количество его «+» зарядов, а, следовательно, и гиперполяризация уменьшается.
В результате возбудимость увеличивается, но она остается ниже, чем в состоянии покоя
Периэлектротон — обратное изменение возбудимости вне электротонических областей.
В зоне прекращения действия катода возбудимость снижается.
В зоне прекращения влияний анода она, наоборот, увеличивается
При длительном прохождении постоянного тока через ткань происходит извращение измененной возбудимости
При длительном замыкании под катодом происходит:
·инактивация Na проницаемости
·повышение уровня критической деполяризации
·увеличение порога деполяризации
·снижение вначале повышенной возбудимости
Это явление получило название катодическая депрессия
При длительном замыкании под анодом происходит:
·снижение К проницаемости
·уровень критической деполяризации снижается
·порог деполяризации уменьшается
·повышается вначале сниженная возбудимость
Это явление получило название анодическая экзальтация
Этот закон необходимо учитывать в медицинской практике
С одной стороны, он может быть использован, если требуется заблокировать проведение возбуждения по нервной или мышечной ткани (катодическая депрессия) или повысить возбудимость (анодическая экзальтация)
С другой стороны, необходимо помнить о возможности привыкания к длительному действию раздражителей, в частности, фармацевтических препаратов, влияющих на возбудимость мембраны.
ФИЗИОЛОГИЯ МЫШЦ И НЕРВОВ
Структура миофибрилл и ее изменения при сокращении.Миофибриллы представляют собой сократительный аппарат мышечного волокна. В поперечнополосатых мышечных волокнахмиофибриллы-разделены на правильно чередующиеся участки (диски), обладающие разными оптическими свойствами. Одни участки анизотропны,т. е. обладают двойным лучепреломлением. В обыкновенном свете они ^выглядят темными, а. в поляризованномг— прозрачными в продольном направлении й непрозрачными в поперечном. Другие участки в Обыкновенной свете выглядят светлыми — они изотропны,т. е. не ; обладают двойным’ лучепреломлением (рис. 34, а) . Анизотропные участки обозначают буквой А,изотропные — буквой I.В середине диска Аразличается светлая полоска Н, посередине диска / — темная «полоска -представляющая собой тонкую мембрану, сквозь поры которой проходят миофибриллы. Благодаря наличию этой опорной структуры параллельно расположенные однозначные диски отдельных фибрилл внутри одного волокна во время сокращения не смещаются по,отношению друг к другу.
Современные представления о структуре миофибриллярного аппарата основываются на исследованиях структуры мышечного волокна-при помощи электронной микроскопии, рентгеноструктурного анализа, фазово-контрастной и интерференционной микроскопии в сочетании с гистохимическими методами.
Установлено, что каждая миофибрилла мышечного волокна диаметром около 1 мкм состоит в среднем из 2500 протофибрилл,представляющих собой удлиненные полимери-, зованные молекулы белков миозина и актина; Миозиновые протофибриллы, или, как, их принято обозначать, нити, вдвое толще актиновых. Их диаметр,примерно 10 нм. В состоянии покоя мышечного волокна нити расположены в миофибрилле таким образом, ^что тонкие длинные актиновые нити входят своими концами в промежутки между толстыми и более короткими миозиновыми нитями (рис. 34, б). Благодаря этому диски / состоят только из актиновых нитей, а диски А — из нитей миозина, а, возможно, еще и другого белка.


Рис. 34. Структура миофибриллы (схема), Показаны диски А и 1-полоски Zи Н(а). Взаимное расположение толстых (миозиновых) и тонких ; (актиновых) нитей в расслабленной (б) й сокращённой (в) миофибрилле.
Светлая полоска Н представляет собой узкую зону, свободную от актиновых нйтей. Мембрана Z,проходя через середину диска /, скрепляет,между собой эти Нити. Важным компонентом ультрамикроскопической-структуры миофибрилл являются также многочисленные поперечные мостики, .соединяющие между собой миозиновые и актиновые нити. При сокращений мышечного волокна указанные нити не укорачиваются, а начинают «скользить» друг по другу: актиновые нити вдвигаются в промежутки между миозиновыми, в результате чего диски / укорачиваются, а диски/1 сохраняют свой размер. Почти исчезает светлая полоска //, так как актиновые нити при сокращении сближаются друг с другом своими концами (рис. 34, в). Причиной «скольжения» является химическое взаимодействие между актином и миозином в присутствии ионов Са 2+ и АТФ. Наблюдается своегорода ч химическое «зубчатое колесо», как 6^,1 протягивающее одну Группу нитей по другой. Роль «зубчиков» в этом процессе приписывают поперечным мостикам, обеспечивающим взаимодействие активных центров белков миозиновых и актиновых нитей.
Это глобулярный белок, состоящий из одного полипептида, который полимеризуется с другими молекулами актина и образует две цепи, обвивающие друг друга (рис. 4-4 А). Такая двойная спираль представляет собой остов тонкого филамента. На каждой молекуле актина есть участок связывания миозина. В покоящемся мышечном волокне взаимодействие между актином и миозином предотвращают два белка — тропонин и тропомиозин (рис. 4-4 Б).

Тропомиозин — стержневидная молекула из двух полипептидов, обвивающихся друг около друга; молекула соответствует в длину примерно семи мономерам актина. Цепи из молекул тропомиозина, уложенные конец в конец, располагаются вдоль всего тонкого филамента. Молекулы тропомиозина частично прикрывают участки связывания каждой молекулы актина, мешая контакту миозина с актином. В таком блокирующем положении молекула тропомиозина удерживается тропонином.
Тропонин — гетеротримерный белок. Он состоит из тропонина Т (отвечает за связывание с одиночной молекулой тропомиозина), тропонина С (связывает ион Са 2+ ) и тропонина I (связывает актин и ингибирует сокращение). Каждая молекула тропомиозина связана с одной гетеротримерной молекулой тропонина, которая регулирует доступ к участкам связывания миозина на семи мономерах актина, прилегающих к молекуле тропомиозина.
Это единое название большой семьи протеинов, имеющих определенные отличия в клетках разных тканей. Миозин присутствует у всех эукариотов. Около 60 лет назад было известно два типа миозина, которые сейчас называют миозин I и миозин II. Миозин II был первым из числа открытых миозинов, и именно он принимает участие в мышечном сокращении. Позднее были открыты миозин I и миозин V (рис. 4-4 В). В последнее время показано, что миозин II участвует в мышечном сокращении, тогда как миозин I и миозин V вовлечены в работу подмембранного (кортикального) цитоскелета. В настоящее время идентифицировано более 10 классов миозина. На рисунке 4-4 Г показано два варианта схемы строения миозина, который состоит из головки, шейки и хвоста. Молекула миозина состоит из двух больших полипептидов (тяжелых цепей) и четырех меньших (легких цепей). Эти полипептиды составляют молекулу с двумя глобулярными «головками», которые содержат оба вида цепей, и длинным стержнем («хвостом») из двух переплетенных тяжелых цепей. Хвост каждой молекулы миозина располагается вдоль оси толстого филамента, а две глобулярные головки выступают по бокам, их иначе называют поперечными мостиками. На каждой глобулярной головке находятся по два участка связывания: для актина и для АТФ. Участки связывания АТФ обладают также свойствами фермента АТФазы, гидролизующей связанную молекулу АТФ.
Источник