Биофизика органов чувств изучает
Название работы: ЭЛЕМЕНТЫ БИОФИЗИКИ ОРГАНОВ ЧУВСТВ
Предметная область: Биология и генетика
Описание: Функцию получения и переработки информации об условиях внешней среды в организме выполняют органы чувств. По существу органы чувств представляют собой измерительные устройства для анализа внешних физических стимулов а также для оценки эффективности действий производимых организмом. Таким образом органы чувств выполняют роль обратной информационной связи в системе организмсреда.
Дата добавления: 2015-05-13
Размер файла: 147.4 KB
Работу скачали: 14 чел.
Организм представляет собой сложную самоорганизующуюся систему, которая характеризуется обменом веществ, энергией и информацией с окружающей средой. Целесообразное реагирование организма, при постоянно меняющихся условиях внешней среды, возможно только при наличии непрерывного поступления в организм информации от окружающей среды. Функцию получения и переработки информации об условиях внешней среды в организме выполняют органы чувств. По существу, органы чувств представляют собой измерительные устройства для анализа внешних физических стимулов, а также для оценки эффективности действий, производимых организмом. Таким образом, органы чувств выполняют роль обратной информационной связи в системе «организм-среда». В 1968 г. Мэчи предложил следующую схему, показывающую взаимодействие организма с внешней средой. Схема имеет вид:
Схема, показывающая взаимодействие организма и внешней среды (по Мэчину)
При исследовании работы органов чувств представляют интерес два аспекта:
Кибернетический аспект заключается в изучении принципов кодирования и передачи информации в органах чувств.
Биофизический аспект заключается в исследовании конкретных физико-химических процессов взаимодействия факторов внешней среды с органами чувств, что приводит к трансформации энергии внешнего воздействия в специфические сигналы, пригодные для анализа нервной системой.
При действии внешнего стимула на органы чувств у человека, возникает специфическое ощущение. Зависимость между величиной ощущения и величиной действующего стимула сформулирована в законе Вебера-Фехнера. В 1760 г. Буггер установил, что соотношение минимального воспринимаемого изменения освещенности к величине освещенности есть величина постоянная в широких пределах освещенности:
Такое же постоянство отношения минимально воспринимаемого прироста раздражения ( ) к его исходной величине ( R ) было установлено Вебером и для других сенсорных систем, то есть:
Так, например, прирост в весе груза, чтобы стать ощутимым, должен превышать ранее действующий вес на 3%. Фехнер предположил, что минимальный прирост ощущения ( ) относительно исходного уровня ощущения ( S ) также постоянен, тогда:
, где k — коэффициент пропорциональности.
Интегрируя это уравнение и проведя алгебраические преобразования, Фехнер получил следующее выражение:
S = a lg R + b , где a , b постоянные.
Таким образом, согласно закону В-Ф, величина ощущения S пропорциональна log силы раздражения. Эта зависимость обусловлена принципом кодирования информации в рецепторном аппарате органов чувств.
Рецепторы представляют собой или специфические окончания афферентных нервных волокон, или нервные окончания в соединении со специализированными клетками, или структурами. Во всех случаях рецептор действует, как преобразователь энергии раздражения в энергию нервного ответа.
При действии внешнего стимула изменяется ионная проницаемость рецепторных мембран, что вызывает их деполяризацию. Это возникает под действием стимула и деполяризация называется генераторным потенциалом (ГП). Величина ГП пропорциональна log интенсивности действующего раздражителя. ГП рецептор клетки генерирует в афферентном нервном волокне ПД, который служит сигналом, передающим информацию в нервной клетке и нервной системе. Между величиной ГП и частотой появления ПД в афферентном волокне, наблюдается линейная зависимость.
Нервное волокно может находиться в двух состояниях:
1 в возбужденном, когда есть ПД;
2 в невозбужденном, когда ПД отсутствует.
Таким образом, в нервной системе имеется дискретная двоичная форма кодирования информации, как в цифровых ЭВМ. Как показывает эксперимент, информация в нервной системе кодируется не последовательностью ПД, а их частотой появления, как в аналоговых вычислительных машинах, где различные модулированные показатели, представленные аналогами, изменяются пропорционально величине модулированных факторов.
Поскольку между интенсивностью стимула и величиной ГП существует логарифмическая зависимость, а ГП связан с частотой ПД линейно, интенсивность стимула и частота ПД должны быть связаны логарифмической функцией. Работы Метьюса, Грэма и других, выполненные с помощью микроэлектронной техники, подтверждают, что частота пропорциональна логарифму величины действующего раздражителя.
Таким образом, по-видимому, данная зависимость преобразования интенсивности действующего стимула ( R ) в частоту нервных импульсов в афферентных волокнах и обуславливает логарифмическую зависимость величины ощущения от интенсивности раздражителя.
ЭЛЕМЕНТЫ БИОФИЗИКИ СЛУХА
ФУНКЦИИ ВСПОМОГАТЕЛЬНЫХ ЭЛЕМЕНТОВ
НАРУЖНОГО И СРЕДНЕГО УХА
В органе слуха принято выделять три части:
- наружное ухо;
- среднее ухо;
- внутреннее ухо.
Упрощенную схему органа слуха можно представить в виде:
Схема органа слуха.
СП слуховой проход; БП барабанная перепонка; М молоточек; Н наковальня;
С стремечко; ОО овальное окно; ВЛ вестибулярная лестница;
УП улитковый проток; БЛ барабанная лестница; Г геликотрема;
КО круглое окно; НУ наружное ухо; СУ среднее ухо.
Слуховые рецепторы расположены во внутреннем ухе, тогда, как вспомогательные элементы, способствующие слуховой рецепции, присутствуют во всех частях органа слуха.
Наружное ухо включает ушную раковину и слуховой проход. С ним связано свойство направленности слухового восприятия, под которым понимают лучшее восприятие звуков, идущих в определенном направлении. Это обеспечивает локализацию источника звука в пространстве (ототолику), кроме этого, слуховой проход служит резонатором. Его резонансная частота у человека лежит в области частот примерно 3 кГц, но резонансная кривая имеет уплощенную форму, поскольку добротность СП, как колебательной системы, невелика. Разница в коэффициенте передачи звуков резонансной и не резонансной частот не более 1 порядка. Границей между наружным и средним ухом служит БП. НУ защищает БП от механических повреждений и, поддерживая определенные микроклиматические условия (температуру, влажность и пр.), стабилизирует ее механические свойства. Таким образом, НУ выполняет следующие функции:
- Локализация звука в пространстве.
- Усиление звукового сигнала (резонансное).
- Механическая защита БП.
- Обеспечение микроклимата, что необходимо для нормального функционирования БП.
По СУ звук проводится цепочкой слуховых косточек: М, Н, С. рукоятка М зафиксирована на БП, а его головка образует сустав с Н, который сочленяется с С. Основание С погружено в ОО, которое служит входом во ВУ, поэтому система БП и слуховых косточек СУ выполняет роль связующего звена между атмосферой и ВУ, заполненным жидкостью. Если бы звук переходил из воздуха в жидкость непосредственно, то интенсивность звуковой волны падала бы из-за сильного отражения звука на границе раздела сред с различным акустическим импедансом [ ]. Жидкость обладает большей по сравнению с воздухом. БП (исключает) и система слуховых косточек обеспечивает согласование воздуха и жидкости ВУ, вследствие чего уменьшаются потери интенсивности звуковой волны при переходе из атмосферы во ВУ.
Согласование и усиление звука обеспечивается благодаря двум обстоятельствам:
- Поверхность БП примерно в 25 раз больше площади ОО, за счет разницы в площадях на входе и выходе системы механической передачи происходит усиление давления на жидкость ВУ, относительно звукового давления, воздействующего на БП.
- Слуховые косточки так расположены в цепочке, что при их движении, они образуют рычаги силы и увеличивают давление на основание С по сравнению с давлением на рукоятку М. Вся система передачи звука в СУ работает наподобие гидравлического пресса с примерно 90 кратным выигрышем в силе.
Максимальный коэффициент усиления давления характерен для звуковых волн с f примерно 1 кГц. Звуковые колебания, как меньших, так и больших f , усиливаются слабее, так как для них слуховые косточки СУ хуже согласовывают , и часть звуковых волн с f примерно 1 кГц, при их средней интенсивности, передается на ВУ практически без потерь. Однако, и для них коэффициент передачи может снижаться и оказывается даже меньше 1. Это имеет место в том случае, когда звук становится слишком интенсивным. Это сопровождается ощущение покалывания, а затем и боли в ушах. Понижение коэффициента передачи происходит при помощи тоненьких мышц, которые прикрепляются к М и С. В ответ на сильные звуки, они рефлекторно сокращаются и уменьшают подвижность системы слуховых косточек.
Итак, систему передачи звука, сосредоточенную в СУ, можно считать механическим преобразователем (усилителем), обладающим переменными регулируемым коэффициентом передачи давления с БП на жидкость ВУ. При разрушении слуховых косточек слух полностью не теряется, но понижается примерно в 1000 раз.
СУ выполняет следующие функции:
- Связующую между атмосферой и ВУ.
- Согласования атмосферы (воздуха) и жидкости ВУ.
- Усиление звукового сигнала.
- Защитная, при больших интенсивностях звука.
Источник
Детальная информация
| Название: | Биофизика сенсорных систем: учебное пособие для студентов высших учебных заведений, обучающихся по направлению подготовки 553100 “Техническая физика” |
| Авторы: | Бигдай Елена Владимировна ; Говардовский Виктор Исаевич ; Лычаков Дмитрий Витальевич ; Самойлов Владимир Олегович |
| Другие авторы: | Самойлов Владимир Олегович |
| Организация: | Санкт-Петербургский государственный политехнический университет |
| Выходные сведения: | Санкт-Петербург: Изд-во Политехн. ун-та, 2005 |
| Электронная публикация: | Санкт-Петербург, 2020 |
| Коллекция: | Учебная и учебно-методическая литература ; Общая коллекция |
| Тематика: | Биофизика ; органы чувств ; учебники и пособия для вузов |
| УДК: | 577.354(075.8) |
| Тип документа: | Учебное издание |
| Тип файла: | |
| Язык: | Русский |
| Код специальности ФГОС: | 16.00.00 |
| Группа специальностей ФГОС: | 160000 — Физико-технические науки и технологии |
| DOI: | 10.18720/SPBPU/2/si20-1511 |
| Права доступа: | Доступ по паролю из сети Интернет (чтение, печать, копирование) |
Группа: Анонимные пользователи
Аннотация
Пособие соответствует авторскому курсу дисциплины «Актуальные проблемы биофизики» направления подготовки 553100 «Техническая физика» по магистерской программе 553120 «Медицинская и биоинженерная физика». Изложены современные представления о биофизических механизмах функционирования сенсорных систем (органов чувств) человека и животных. Рассмотрены общие принципы преобразования рецепторами сигналов внешней среды в нервную импульсацию. Приведены сведения о трансдукции в зрительной, слуховой, вестибулярной, интероцептивной, вкусовой и обонятельной сенсорных системах. Предназначено для магистров факультета медицинской физики и биоинженерии СПбГПУ, может быть рекомендовано студентам других факультетов, изучающим основы биологии и медицины.
Печатается по решению редакционно-издательского совета Санкт-Петербургского государственного политехнического университета.
Права на использование объекта хранения
| Место доступа | Группа пользователей | Действие |
|---|---|---|
| Локальная сеть ИБК СПбПУ | Все |    |
| Интернет | Авторизованные пользователи | |
 »/> »/> | Интернет | Анонимные пользователи |
Оглавление
- ОГЛАВЛЕНИЕ
- Введение
- 1. Механизмы преобразования информации в рецепторах сенсорныхсистем
- 2. Биофизика фоторецепции (механизмы фототрансдукции в фоторецепторах позвоночных)
- 3. Топография органов, расположенных в перепончатом лабиринте височной кости
- 4. Биофизика слуха
- 5. Биофизика вестибулярного аппарата
- 6. Биофизика хемосенсорных систем
- Библиографический список
Статистика использования
| Количество обращений: 2 За последние 30 дней: 0 Подробная статистика |
Сайт и управление доступом на базе АБИС «Руслан-Нео» (© ООО «ОБС»)
Источник
Глава 13 ЭЛЕМЕНТЫ БИОФИЗИКИ ОРГАНОВ ЧУВСТВ
ОБЩИЕ ЗАКОНОМЕРНОСТИ
Организм представляет собой сложную саморегулирующуюся систему, которая характеризуется обменом веществ, энергии и информации с окружающей средой.
Целесообразное реагирование организма при постоянно меняющихся условиях внешней среды возможно только при наличии непрерывного поступления в организм информации от этой среды. Функцию получения и переработки информации об условиях внешней среды в организме выполняют органы чувств. По существу органы чувств представляют собой измерительные устройства для анализа внешних физических стимулов, а также для оценки эффективности действий, произведенных организмом. Таким образом, органы чувств выполняют роль обратной информационной связи в системе организм— среда. На рис. 55 изображена схема, показывающая роль органов чувств как обратной связи в системе организм — среда.
При исследовании работы органов чувств представляют интерес два аспекта: кибернетический и биофизический. Кибернетический аспект заключается в изучении принципов кодирования и переработки информации в органах чувств, а биофизический — в исследовании конкретных физико-химических процессов взаимодейст-

вия факторов внешней среды с органами чувств, приводящих к трансформации энергии внешнего воздействия в специфические сигналы, пригодные для анализа нервной системой.
При действии внешнего стимула на орган чувств у человека возникает специфическое ощущение. Зависимость между величиной ощущения и величиной действующего стимула сформулирована в законе Вебера — Фехнера. В 1760 г. Буггер установил, что отношение минимально воспринимаемого изменения освещенности А/ к величине освещенности I есть величина постоянная в шиооких пределах изменения освещенности:

(1)
Такое же постоянство отношения минимально воспринимаемого прироста раздражения ΔR к его исходной величине Rбыло установлено Вебером для других сенсорных систем (мышечно-суставной, слуховой и др.):
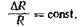
Например, прирост в весе груза, чтобы стать ощутимым, должен превышать ранее действовавший вес на 3%. Фехнер предположил, что минимальный прирост ощущения А5 относительно исходного уровня ощущения S тоже константа, т. е.

(2>
где k — коэффициент пропорциональности. Интегрируя уравнение (2) и производя алгебраические преобразования, Фехиер получил:
(3)
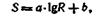
где а и Ь — постоянные величины. Таким образом, согласно закону Вебера — Фехнера, величина ощущения 8 возрастает пропорционально логарифму силы раздражения. Эта зависимость обусловлена принципом кодирования информации в рецепторном аппарате органов чувств.
Рецепторы представляют собой или специализированные окончания афферентных нервных волокон, или нервные окончания в соединении со специализированными клетками или структурами. Во всех случаях рецептор действует как преобразователь энергии раздражите-теля в энергию нервного ответа.
При действии внешнего стимула изменяется ионная проницаемость рецепторных мембран, что вызывает их деполяризацию. Эта возникающая под действием стимула деполяризация называется генераторным потенциалом. Рецепторные клетки не подчиняются закону «все или ничего» — величина генераторного потенциала пропорциональна логарифму интенсивности действующего раздражителя. Генераторный потенциал рецептор-ной клетки генерирует в афферентном нервном волокне потенциалы действия (ПД), которые служат сигналами, передающими информацию в нервной клетке и нервной системе. Между величиной генераторного потенциала и частотой появления ПД в афферентном волокне наблюдается линейная зависимость.
Нервное волокно может находиться в двух состояниях: возбужденном — есть ПД, и невозбужденном — ПД отсутствует. Таким образом, в нервной системе имеется дискретная двоичная форма кодирования информации. Такая форма кодирования применяется в цифровых вычислительных машинах, где вся информация кодируется двумя цифрами: 1 и 0. Однако принцип кодирования информации в нервной системе отличается от такового в цифровых машинах. Как показывают эксперименты, информация в нервной системе кодируется не последовательностью ПД, как в цифровых машинах (последовательностью единиц и нулей), а частотой появления ПД, как в аналоговых вычислительных машинах, где различные моделируемые показатели представлены аналогами, изменяющимися пропорционально величине моделируемых факторов.
Поскольку между интенсивностью стимула и величиной генераторного потенциала существует логарифми-
ческая зависимость, а генераторный потенциал связан с частотой ПД линейно, то интенсивность стимула и частота ПД должны быть связаны логарифмической функцией. Работы Мэтьюса, Хартлайна, Грэма, выполненные с помощью микроэлектродной техники, подтвердили, что частота fпоявления ПД в афферентных нервных волокнах пропорциональна логарифму величины действующего раздражителя:

где т и п — постоянные величины.
По-видимому, данная зависимость преобразования интенсивности действующего стимула в частоту нервных импульсов в афферентных волокнах и обусловливает логарифмическую зависимость величины ощущения от интенсивности раздражителя.
ВОСПРИЯТИЕ ВКУСА
В эволюции животных огромное значение имело контактное, дифференцированное распознавание пищи, в результате которого происходила или ее утилизация, или, напротив, отклонение.
Химические соединения, входящие в состав пищи животных, действуя на вкусовые рецепторы, обусловливают ее соленый, кислый, сладкий или горький вкус. Имеются более сложные системы классификации вкусовых ощущений, однако для человека, вероятно, справедливо положение о наличии четырех элементарных вкусовых ощущений.
У человека вкусовые луковицы, содержащие вкусовые рецепторы, находятся во вкусовых сосочках, расположенных на поверхности языка, на задней стенке глотки, мягком небе, миндалинах и надгортаннике. Вкусовые луковицы, имеющие овальную форму, содержат вкусовые и опорные клетки и сообщаются с поверхностью слизистой оболочки вкусовыми порами. Раньше считали, что опорные клетки не имеют отношения к вкусовой рецепции; в настоящее время многие исследователи рассматривают опорные клетки в качестве переходной стадии дифференцирующихся рецепторных.
Рецепторные клетки в апикальной области имеют микровиллы (микроворсинки), которые в виде кисточки
через вкусовые поры сообщаются с жидкой средой, покрывающей слизистую оболочку. Предполагают, что на поверхности микровилл расположены активные белковые центры, с которыми реагируют молекулы или ионы вкусовых веществ. Пространство между микровиллами заполнено особой субстанцией мукополисахаридной в мукопротеидной природы. Некоторые авторы считают, что эта субстанция функционирует наподобие ионообменной смолы, давая возможность молекулам или ионам одних веществ контактировать с поверхностью микровилл, а других — нет. У основания луковицы вкусовые клетки образуют синапсы с окончаниями афферентных нервных волокон лицевого, языкоглоточного и блуждающего нервов.
В последнее десятилетие были проведены большие исследования вкусового анализатора с помощью микроэлектродной техники. Было установлено, что вкусовые клетки имеют потенциал покоя, величина которого колеблется в пределах 30—50 мВ. При действии вкусового стимула на клетку наблюдается медленная деполяризация клеточной мембраны — формируется генераторный потенциал. Его величина в зависимости от интенсивности стимула варьирует в пределах от 15 до 45 мВ.
Механизм передачи возбуждения от вкусовой клетки к нервному волокну окончательно не выяснен, однако известно, что передача в синапсе осуществляется при участии ацетилхолина.
Оказалось, что одна и та же клетка может реагировать на сладкие, соленые и кислые стимулы, тогда как другие клетки обладают повышенной чувствительностью только к одному или двум стимулам. В отличие от вкусовых клеток вкусовые сосочки строго специфичны. При электрическом раздражении отдельных сосочков возникает какое-либо одно из четырех вкусовых ощущений. Химическая стимуляция этих же сосочков у человека вызывала аналогичные ощущения. На основе подобных опытов Бекеши (1966) пришел к заключению, что существует полное совпадение вкусовых качеств для каждого отдельного сосочка, стимулированного как электрическим, так и химическим путем. Сосочки отличаются строгой локализацией на поверхности языка и делятся на соленые, кислые, сладкие и горькие. Возможно, что-строгая специфичность сосочков обусловлена ионообмен-
ными свойствами субстанции, заполняющей пространство между микровиллами и обусловливающей избирательную проницаемость вкусовых пор для вкусовых веществ.
В опытах с отведением потенциалов от афферентных нервов было обнаружено, что небольшое количество нервных волокон обладает узкой вкусовой специфичностью, а большая часть волокон обладает или относительной специфичностью, или вообще не обладает специфичностью. Следует полагать, что наличие специфичности в периферическом отделе вкусового анализатора обусловливает формирование дифференцированных вкусовых ощущений в центральном отделе. Натуральное пищевое раздражение одновременно возбуждает несколько или все вкусовоспринимающие элементы, в результате чего возникает сложное, смешанное вкусовое ощущение.
Интересно отметить, что у мухи удалось выделить три узкоспецифичных хеморецептора: один из них стимулируется растворами солей, другой — растворами сахара, глюкозы и т. д., третий стимулируется воздействием чистой воды. У человека чувствительности к чистой воде не обнаружено.
Химические свойства вкусовых веществ и теории вкуса
Все вкусовые вещества делятся на три группы:
1) вещества, близкие по своей химической структуре и вызывающие примерно одинаковые вкусовые ощущения;
2) вещества, близкие по химическим свойствам, но обладающие разным вкусом; 3) вещества разной структуры, но сходного вкуса. К первой группе относятся сахара: глюкоза, сахароза, галактоза, лактоза. Они обладают сладким вкусом. Кислым вкусом обладают диссоциированные неорганические и органические кислоты. Ко второй группе можно отнести ряд изомеров. Например, d-валин имеет горьковато-сладкий вкус, а l-валин — сладкий вкус, d-лейцин — сладкий, l-лейцин — горький. К третьей группе можно отнести, например, полисахариды, глицерин, гликокол, сахарин, нитробензол, хлористый метил, хлороформ, резорцин, имеющие сладкий вкус, а также дифенил-мочевину, стрихнин, хинин, ни-
тропроизводные бензола, многие неорганические соли и кислоты, имеющие горький вкус. Аналогичность вкуса при таком несходстве химической структуры вещества относится главным образом к сладкому и горькому вкусу.
Какие же свойства вкусовых веществ обусловливают их вкус? Соленый вкус NaCl присущ и другим хлоридам (К, NH3, Са и т. д.). Можно было бы предположить, что свободный анион С1 — , а не катионы Na + , К + , NН4, Са ++ ответствен за соленый вкус. Однако значение катионов тоже нельзя исключить. Так, например, в слабой концентрации NаС1 обладает соленым вкусом, NаВr — горьковато-соленым, Nа1 — солоновато-горьким. Можно заключить, что ион N + придает растворам всех этих солей присущий им солоноватый вкус. В настоящее время считают, что в основе соленого стимула лежит действие низкомолекулярных анионов и катионов. Принимают, что интенсивность вкуса низкомолекулярных солей зависит от катионов Nа + , К + , Rb + , Сs+ и т. д., а характер вкуса — от анионов С1 -, Br — , I — и т. д.
Кислым вкусом обладают большинство органических и неорганических кислот. Как известно, общая черта всех кислот — способность к диссоциации в водных растворах на анионы и катион водорода. Кислый стимул связывают с воздействием на вкусовые клетки водородных ионов. В этом убедились при помощи довольно простых опытов. Например, НС1 ощущается кислой при разведении N/800, в то время как при этой же концентрации практически весь диссоциированный NaС1 безвкусен. Воздействие Н + пропорционально его концентрации. Однако более сильные кислоты в одинаковой концентрации со слабыми не всегда вызывают более сильное вкусовое ощущение. Это объясняется тем, что для ионов Н + проницаемость субстанции, окружающей микровиллы вкусовых клеток, имеет низкое значение. Слабые же кислоты в виде недиссоциированных молекул достигают микровилл вкусовых клеток и затем диссоциируют; образовавшиеся ионы Н + взаимодействуют с активными центрами микровилл.
Сладкий вкус вызывается большим числом органических веществ, которые, как правило, не диссоциируют. В настоящее время окончательно не выяснено, какие свойства молекул вызывают сладкий вкус. Многие сладкие вещества являются многоатомными спиртами с по-
19 Медицинская биофизика
вторяющейся —СН2ОН группой (сахара, глицерин). Сахарин, который также обладает сладким вкусом, имеет совершенно другую структуру. По мнению Акри, сладкий стимул молекул связан с их возможностью образовывать слабые водородные связи с активным центром «рецепторной» молекулы, локализованной во вкусовой клетке. Во всех случаях структура молекулы должна соответствовать стерическому положению и возможностям водородных связей рецепторной молекулы.
Менее всего изучена природа горького стимула вкусовых веществ. Было отмечено, что соли по мере увеличения молекулярного веса становятся все более горькими. Если NаС1 имеет соленый вкус, то СsС1 — горький,-КI тоже часто является горьким. Другие хорошо известные горькие вещества — это свободные основания, например алкалоиды. Имеются данные, что горький стимул молекул, как и сладкий стимул, связан с возможностью образования слабых дисперсионных связей типа водородных.
Существует несколько теорий, пытающихся объяснить конкретные процессы взаимодействия молекул вкусовых веществ с рецепторными клетками. П. П. Лазарев еще в 1920 г., исходя из роли ионов в процессе возбуждения, допускал, что вкусовые луковицы каждого сосочка содержат высокочувствительные вещества белковой природы, разлагающиеся под влиянием адекватного стимула, а ионизированные продукты распада возбуждают соответствующие нервные окончания.
В настоящее время наиболее разработанной является теория Бейдлера. Он предположил, что вкусовые стимулы— молекулы или ионы — взаимодействуют с определенными участками плазматической мембраны вкусовой клетки, которые он выделяет в качестве активных центров. Активные центры представляют собой полиэлектролиты белковой природы, содержащие большое количество заряженных боковых цепей. Взаимодействие вкусовых стимулов с активными центрами протекает по типу мономолекулярной реакции:
где С — концентрация стимулирующего вкусового вещества; N — общее число активных центров рецептора; Z, — число активных центров, связанных при концентрации вкусового вещества, равной С.
Согласно закону действующих масс, константа равновесия К данной реакции будет

(6
Величина ответа R рецепторной клетки прямо пропорциональна числу связанных активных центров (R=Z), а максимальный ответ Rм наблюдается, когда заняты все активные центры (RМ=N). С учетом этих положений уравнение (6) приобретает следующий вид:
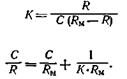
(8)
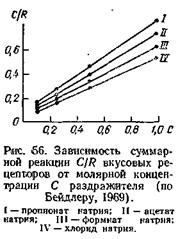
Уравнение (8) представляет собой основное уравнение вкусовой рецепции. Величина С/ R выражает суммарную реакцию вкусовых рецепторов. Согласно уравнению (8), между С/ R и С существует линейная зависимость. Данное положение получило экспериментальное подтверждение (рис. 56).
Бейдлер считает, что взаимодействие частиц вкусового вещества с активными центрами рецептора представляет собой физический процесс адсорбции, в основе которого лежит слабое дисперсионное взаимодействие. Это согласуется с ранее описанным предположением Акри об образовании водородных связей между молекулами сладкого стимула и рецептором.
На основе экспериментальных данных из уравнения (8) или из графиков на рис. 56 можно найти величину константы равновесия данной реакции. Она варьирует от 7,7 для бутирата натрия до 9,8 для хлорида натрия. Эти величины К очень близки к К для адсорбции солей на белках, что соответствует исходному допущению Бейдлера об адсорбции вкусового вещества на поверхности рецептора и о неферментативной природе реакции.
О физической природе процесса свидетельствует также и то, что реакция вкусовых рецепторов на соленый стимул не изменяется при повышении температуры от 20 до 30 °С. Кроме того, величина реакции заметно не изменяется при сдвиге рН от 3 до 11, что исключает воз-
можность сильного электростатического взаимодействия с ионогенными группами белковой молекулы.
Впоследствии термодина
мические расчеты показали,
что этот процесс адсорбции
частиц приводит либо к
уменьшению количества
гидратационной воды на поверхности рецептора, либо к небольшому изменению конформации молекулы рецептора.
Возможно, что изменение конформации рецепторных молекул лежит в основе
дальнейшего процесса, приводящего в конечном счете к возбуждению вкусовой клетки.
Дальнейшее развитие теории вкусового восприятия было связано с открытием Дастоли сладкочувствитель-ных и горькочувствительных белков во вкусовых луковицах млекопитающих. Дастоли удалось выделить из специфических сладкочувствительных и горькочувствительных сосочков белки, избирательно взаимодействующие со сладкими или горькими веществами. Термодинамические и кинетические особенности взаимодействия сладких и горьких веществ с молекулами выделенных белков совпадают с особенностями мономолекулярных реакций, описанных Бейдлером. Поэтому в настоящее время считают, что именно этими белками представлены активные центры вкусовых клеток. Однако локализация этих белков во вкусовых клетках еще не установлена.
Эта теория подтверждает справедливость ранее выдвинутого предположения П. П. Лазарева о наличии нескольких видов вкусочувствительных белков.
По-видимому, следующая задача биофизики вкусового восприятия заключается в исследовании молекулярной структуры выделенных белков. Это позволит проследить шаг за шагом, каким образом возникающие в них конформационные сдвиги приводят к возбуждению рецепторной клетки и его последующей передаче в центральную нервную систему.
ВОСПРИЯТИЕ ЗАПАХА
В основе обоняния лежит взаимодействие молекул пахучих веществ с обонятельными клетками. Молекулы пахучих веществ, отделяясь от своей основной массы и передвигаясь токами воздуха, могут действовать на расстоянии.
Обонятельные рецепторы входят в состав обонятельного эпителия, расположенного в задней части носовой перегородки и в верхнем носовом ходе. Рецепторные клетки представляют собой биполярные нейроны диаметром около 5—10 мкм, расположенные вокруг цилиндрических опорных клеток. У человека количество обонятельных рецепторов достигает 60 млн., а площадь обонятельного эпителия — 5 см 2 . Поверхность обонятельного эпителия покрыта водянистой средой. Периферические отростки обонятельных клеток заканчиваются на своей вершине утолщением грушевидной формы — обонятельной булавой. Как показали электронно-микроскопические исследования, на поверхности каждой обонятельной булавы расположено 9—16 очень тонких ресничек. Центральные отростки отходят от противоположного конца обонятельных клеток и образуют тонкие нити, которые вступают в полость черепа. На нижней поверхности лобной доли они сходятся, образуя обонятельную луковицу, в клубочках которой и заканчивается первый нейрон обонятельного афферентного пути.
При действии обонятельных стимулов на обонятельные рецепторы происходит формирование генераторных потенциалов, вызывающих в центральных отростках рецепторов нервные импульсы.
Суммарный генераторный потенциал обонятельных рецепторов — электроольфактограмму — можно зарегистрировать с помощью электрода, приложенного к поверхности обонятельного эпителия.
Вопрос об избирательности обонятельных рецепторов к молекулам пахучих веществ окончательно не выяснен. До настоящего времени не было обнаружено строгой избирательности рецепторов. Так, некоторыми учеными с помощью микроэлектродов регистрировались потенциалы одиночных волокон обонятельного нерва лягушки при действии различных пахучих веществ. Все исследованные рецепторные нейроны обнаружили грубую избирательную чувствительность к запахам. Каждый из
293
них отвечал на одни запахи и не отвечал на другие. Большинство из них дает особенно сильный ответ по крайней мере на один из 25 использованных запахов и более слабо отвечает на остальные.
В основе возникновения обонятельного ощущения лежит взаимодействие молекул или частиц пахучего вещества с обонятельными рецепторами. Однако до настоящего времени не ясно, какие свойства молекул обусловливают это взаимодействие. Не ясно также, с молекулами каких веществ, локализованных в рецепторных клетках, происходит взаимодействие.
Следует признать, что работа органа обоняния по сравнению с другими органами чувств раскрыта в меньшей степени.
Попытки объяснить пахучие свойства молекул их химическими свойствами и структурой не увенчались успехом. Вещества со сходными свойствами и структурой могут обладать разными запахами, и наоборот. В настоящее время из всех теорий восприятия запаха наибольшего внимания заслуживают две: квантовая и стереохимическая.
Согласно квантовой теории запаха молекулам пахучего вещества свойственны внутримолекулярные колебания, в результате которых они испускают электромагнитное излучение где-то в инфракрасной области спектра. Это излучение взаимодействует с молекулами «обонятельного пигмента» типа каротиноидов, находящегося в мембране рецепторов, что в конечном счете приводит к деполяризации мембраны рецептора. По мнению авторов данной теории, молекулы веществ с похожими запахами должны характеризоваться сходными низкочастотными колебаниями. В качестве подтверждения Райт приводит вещества, обладающие миндальным запахом и близкой частотой колебаний: нитробензол, бензонитрил, бутиронитрил и др.
Из обонятельных клеток удалось выделить ряд каротиноидов и витамин Л, которые, по мнению Райта, поглощают электромагнитное излучение молекул пахучих веществ.
Однако, несмотря на известные успехи, данная теория встречает ряд очень серьезных возражений. Так,
294
многие вещества со сходными запахами имеют различные частоты колебаний и разные спектры поглощения в инфракрасной области, например многие спирты. Молекулы, в которых атом водорода замещен изотопом дейтерия, обладают сходным запахом, хотя при этом значительно меняются частоты основных колебаний. С другой стороны, молекулы с почти одинаковыми частотами вибрации и спектрами инфракрасного поглощения обладают несходными запахами, например d— и l- изомеры. Кроме того, не установлена локализация пигментов в обонятельных клетках и не доказано их участие в первичном процессе взаимодействия с пахучими молекулами.
Более обоснованной является стереохимическая теория восприятия запаха, выдвинутая Монкрифом и подробно разработанная Эймуром в 1962 г. Согласно этой теории, запах вещества обусловлен не химическим составом молекул, а их формой и размерами, Обонятельная система состоит из рецепторов разных типов, каждый из которых соответствует отдельному первичному запаху. Поверхности рецепторных клеток имеют углубления— лунки — определенной формы. Молекулы пахучего вещества вызывают ощущение запаха в том случае, если их форма соответствует форме лунок рецептора, в которые они плотно входят. Этот принцип аналогичен принципу «замка и ключа» при взаимодействии фермента и субстрата. Согласно представлениям Эймура, вещества, имеющие сходный запах, должны также обладать сходной формой своих молекул,
Эймур, сопоставив большое количество данных о форме молекул с ощущениями, которые они вызывают, пришел к выводу о наличии семи первичных (простых) запахов: камфароподобного, мускусного, цветочного, мятного, эфирного, острого и гнилостного. В этом смысле первичные запахи аналогичны трем основным цветам зрительного и четырем вкусам вкусового анализаторов. Каждому первичному запаху соответствует определенная форма молекул и лунки на поверхности рецептора. Так, эфирная молекула отличается палочковидной формой. Она должна иметь вытянутую лунку: ее длина равна 1,8 нм, ширина 0,5 нм, глубина—-0,4 нм. Камфорная молекула имеет сферическую форму диаметром 0,7 нм, а ее лунка имеет форму эллиптической чаши глубиной 0,4 нм, длиной 0,9 нм и шириной 0,75 нм. Молекулы, вы-
зывающие другие ощущения, имеют более сложную форму. Если сложная молекула внедряется сразу в две лунки, то возникает сложный запах.
Теория Эймура нашла свое подтверждение в том, что в некоторых случаях оказалось возможным предсказать запах веществ, исходя из формы их молекул. Так, было синтезировано несколько органических веществ, рассчитана их вероятная форма и предсказан их запах.
Однако данная теория также имеет ряд недостатков. Например, острые и гнилостные запахи не вмещаются в стереохимическую схему. Для молекул, вызывающих острые и гнилостные запахи, форма и размер не имеют значения; решающую роль играет электрический заряд. Острые едкие запахи свойственны тем соединениям, молекулы которых из-за нехватки электронов имеют положительный заряд и сильное сродство к электронам. Гнилостные запахи, наоборот, вызываются молекулами, обладающими избытком электронов.
Кроме того, данная теория обходит молчанием вопрос о том, что происходит после попадания молекулы в соответствующую лунку на поверхности рецептора, с чем она в ней взаимодействует при этом. Теория не объясняет, как возникают потенциалы в ответ на заполнение лунок и т. д.
Итак, заканчивая рассмотрение этого вопроса, приходится признать, что нам все еще не хватает каких-то важных сведений о свойствах пахучих молекул. По-видимому, имеют значение как их структура и форма, так и квантовые свойства. Но они должны обладать еще какими-то, пока неизвестными, универсальными свойствами, которые и обусловливают их запах.
СЛУХ
Из всех органов чувств наиболее сложные преобразования при восприятии стимула наблюдаются в органе слуха. Восприятие звука должно рассматриваться как достигшая-надбальшего развития механорецепция. С по-мощю системы механорецепторов мозг получает инфор-мацию охарактере субстрата, с которым контактирует тело,о скорости тока вды, ветра, о давлении жидкости в сосудах и полостях тела (прессорецепция), о степени растяжения. мышц сухожилий(проприорецепция), о

изменении гравитационного поля, ускорениях к вибрациях тела и, наконец, о наличии звуковых колебаний.
Периферический отдел слухового анализатора включает в свой состав наружное, среднее и внутреннее ухо (рис. 57). Наружное ухо состоит из ушной раковины и наружного слухового прохода.. Он заканчивается барабанной перепонкой, отделяющей наружное хо от среднего. Существеннейшей частью среднего уха является цепь косточек —молоточек, Наковальня и стремечко, которые передают колебания барабанной перепонки внутреннему уху этих косточек — молоточек — вплетена сваей рукояткой в барабанную перепонку, другая сторона молоточка сочленена с наковальней, которая в свою очередь связана со стремечком.
Во внутреннем ухе находится улитка, которая является вопринимающей ‘частью слухового анализатора. Полость улитки делится двумя мембранами — основной и тонкой вестибулярной — на три канала. Верхний и нижний каналы заполнеы жидкостью —перилимфой — и сообщаются между собой у вершины улитки через небольшое отверттие — геликотрему. Верхний канал отделен от полости среднего уха мембраной овального окна, к которому прикреплено стремечко, а нижний канал —
мембраной круглого окна. Средний канал заполнен эндолимфой.
Внутри среднего канала улитки на основной мембране расположен звукопринимающий аппарат — корти
‘ев орган, содержещий рецепротные волосковые клетки.
Рецепторные клетки расположены на основной мембране двумя слоями. Внутренний слой образован одним рядом рецепторных клеток, а наружный слои —— тремя четыря рядами клоеток. Рецепторные клетки имеют вытянутую форму и одним концом фиксированы на основной мембране, а другим выходят в полость среднего канала улитки. На этом конце рецепторных клеток имеется по 60—70 волосков длиной несколько микрометров. Над волосковыми клетками по всему ходу канала расположена покровная пластинка. Волоски рецепторных клеток прикреплены к покровной пластинке. Каждая волосковая клетка имеет синапсы с несколькими окончаниями слуховых нейронов.
Звук представляет собой упругие механические колебания среды, распространяющиеся в виде продольных волн сжатия и растяжения. Звук распространяется во всех средах. Человек способен воспринимать звуковые колебания в диапазоне от 20 Гц до 20 кГц.
Звуковые колебанния распространяются по наружному слуховому проходу и вызывают колебание барабанной перепонки. Барабанная перепонка в силу своей геометрии и механических свойств не имеет собственного периода колебаний и колеблется всегда с частотой звуковых колебаний, однако наибольшей чувствительностью обладает к частоте 1000 Гц. Колебания барабанной перепонки через косточки среднего уха передаются мембране овального окна. В результате различной длинны плеч рычагов косточек, а также в результате неравенства площадей барабанной перепонки и овального окна сила колебании мембраны овального окна возрастает приблизительно в 22 раза по сравнению с силой колебаний барабанной перепонки.
Колебания мембраны овального окна вызывают колебания перилимфы в верхнем и нижнем каналах улитки. Вестибулярная мембрана представляет собой очень тонкуюпленку, поэтому колебания перилимфы передаются на эндолимфу и на основную мембрану. При колебании основной мембраны в волосковых клетка возникают генераторные потенциалы, генерирующие нервные
импульсы, которые передаются через синапсы на нервные окончания. До настоящего времени окончательно не выяснено, передается возбуждение в синапсах химическим или электрическим путем; возможно, имеют место оба этих механизма. Так, наличие холинергической системы в волосковых клетках указывает на возможность химической передачи возбуждения, а практическое отсутствие синаптической задержки между действием стимула и тоном действия в слуховом нерве указывает на возможность прямой электрической передачи возбуждения.
Суммарный рецепторный потенциал волосковых клеток — так называемый микрофонный потенциал — можно зарегистрировать, если один электрод ввести в полость улитки, а второй сделать индифферентным. Изменение микрофонного потенциала в точности соответствует воздействующему звуковому стимулу. Частота колебаний микрофонного потенциала соответствует частоте звуковых колебаний, а амплитуда в широких границах пропорциональна интенсивности звука. Если отводимую разность потенциалов усилить и направить в громкоговоритель, то можно получить тот же звук, который воздействовал на ухо. Этот эффект назван микрофонным эффектом улитки.
В настоящее время существуют две основные теории, объясняющие механизм возникновения генераторных потенциалов волосковых клеток: механоэлектрическая (Дэвис, 1957) и цитохимическая (Я. А. Винников, Л. К. Титова, 1961).
При воздействии звуковых колебаний основная мем-брана с прилежащими к ней структурами приходит в колебательное движение. Покровная пластинка прикреплена к кортиеву органу у его внутреннего и внешнего краев, и ее ось вращения расположена значительно выше, чем ось вращения основной мембраны. Поэтому во время колебаний между основной мембраной и покровной пластинкой, к которой прикреплены волоски рецеп-торных клеток, возникают режущие силы и происходит скольжение. Так, как волоски рецепторных клеток прикреплены к покровной пластине, то при ее скольжении происходит сгибание волосков. Согласно механоэлектри-ческой теории, сгибание волосков приводит к изменению ионной проницаемости мембран рецепторных «клеток и к возникновению генераторных потенциалов. Со-
гласно цитохимической теории, при движении основной мембраны и покровной пластинки в области волосков рецепторных клеток появляется ацетилхолин, который вызывает увеличение проницаемости рецепторных мембран для ионов и их деполяризацию. Как следует из изложенного, рассмотренные теории не столько противоречат, сколько дополняют друг друга.
Согласно расчетам Бекеши, электрическая энергия, созданная колебаниями кортиевого органа, значительно превышает поглощенную звуковую энергию. Следовательно, энергия звукового стимула используется лишь для запуска энергетических процессов в рецепторах путем изменения проницаемости мембран. Эти процессы возможны благодаря наличию ионных градиентов в рецепторных клетках. Энергия звукового стимула не только запускает внутренний источник энергии рецепторов, но и модулирует (определяет) его, что подтверждается наличием ранее описанного микрофонного эффекта улитки.
Источник